Зависимость обшей минерализации костей сеголетков карпа от интенсивности их роста, % золы в сухом веществе жаберной крышки
| Сеголетки | Масса рыбы, г | ||||
| 5,2 | 15,8 | 27,4 | 34,7 | 51,3 | |
| Чешуйчатый карп | 23,7 | 33,9 | 48,5 | 54,6 | 56,7 |
| Голый карп | 24,1 | 36,0 | 50,0 | 55,3 | 56,2 |
Примечание. Средние данные по трем водоемам Московской, Смоленской областей и Ставропольского края {октябрь 1983 г.).
Кроме возраста на минерализацию костей влияет видовая принадлежность. У одновозрастных особей карпа, плотвы, окуня и сома из одного водоема различия в степени минерализации жаберной крышки достигают 15 %.
Степень минерализации воды (58-260 мг/л) и характер питания (включая 30-дневное голодание) не влияют на уровень золы в костях рыб. Однако темп роста существенно влияет на этот показатель. Сеголетки карпа, выращенные в одних и тех же условиях, но различающиеся по массе тела, имеют большие различия в степени минерализации костной ткани.
Элементный состав костной золы менее стабилен по сравнению с общей минерализацией и изменяется под влиянием условий содержания рыбы. Для сеголетков карпа разных породных линий (голый, зеркальный, линейный и чешуйчатый) можно привести следующие усредненные характеристики макро- и микроминерального состава костной ткани (табл. 5.2).
Содержание макро-(%) и микроэлементов (мг%) в костной золе сеголетков карпа
| Зола | Са | Р | Са:Р | Mg | Сu | Мn | Zn | Fe |
| 45,3 | 75,0 | 9,3 | 0,220 | 2,41 | 11,39 | 78,57 | 16,75 |
Содержание кальция в сумме макроэлементов велико, однако подвержено большим изменениям, так как кости представляют собой депо этого элемента. В экстремальных условиях уровень кальция в скелете может уменьшаться на 30-35 % без гибельных последствий.
Значительная доля минеральных образований кости представлена соединениями фосфора, входящими в состав гидрооксиапатита. Содержание фосфора в костях рыб в 2 раза ниже по сравнению с наземными животными, но довольно стабильно (около 10 %). Соотношение Са : Р в костях сеголетков карпа со-ставляет примерно 2,7 : 1.
Магний в составе кристаллов гидрооксиапатита обеспечивает прочность костной ткани наземных животных. У рыб требования к прочности костей иные, поэтому уровень магния в костях невысок (220 мг% вместо 1500 мг% у наземных животных). У рыб больше и соотношение Са: Mg (114: 1 у сеголетков карпа и 50 :1 у наземных домашних животных).
Микроминералъный состав костей не отличается единообразием. На него влияют многие факторы (питание, возраст, видовая принадлежность). Однако главным фактором следует считать алиментарный. Соотношение же отдельных микроэлементов в костной ткани при стабильных условиях выращивания рыбы более постоянно. Так, больше всего в костях цинка (60-100 мг% на золу), второе место занимает железо (15-20мт%), далее марганец (7- 16 мг%) и медь (1-5 мг%). Интересно, что концентрация железа в воде не влияет на накопление элемента в скелете.
Концентрация тяжелых металлов в костях напрямую определяйся их распространенностью во внешней среде. Интенсивность аккумуляции тяжелых металлов выше у молоди. Концентрация стронция (Sr90) в костях ушастого окуня и тиляпии может превышать его уровень в воде в 10 раз. У тиляпии уже через 2 дня после содержания ее в радиоактивной воде уровень радиации костей достигает уровня радиации воды. Через 2 мес концентрация стронция в скелете тиляпии в 6 раз превышала таковую в воде. Причем насколько легко тяжелые металлы проникают в костную ткань рыб, настолько же медленно ее покидают. Стронций остается в скелете рыб десятилетиями даже при условии содержания рыбы в свободной от этого элемента среде.
Скелет костистых рыб принято делить на осевой и периферический (см. рис. 5.2).
Осевой скелет включает в себя позвоночный столб (туловищная и хвостовая части), ребра и кости головы. Количество позвонков у разных видов неодинаково и колеблется от 17 у луны-рыбы до 114 у речного угря. У хрящевой рыбы - морской лисицы - количество позвонков достигает 365. Первые четыре туловищных позвонка могут быть трансформированы в так называемый Веберов аппарат.
Позвонки туловищной и хвостовой частей неодинаковы по строению. Туловищный позвонок имеет тело, верхний остистый отросток и два нижних остистых отростка. У основания верхнего остистого отростка и верхнего края тела позвонка находится невральная дуга. Внизу справа и слева от туловищных позвонков отходят ребра, которые соединены с позвонками подвижно.
Позвонки хвостового стебля отличаются тем, что их нижние остистые отростки, срастаясь, формируют гемальную дугу и непарный гемальный отросток. К тому же в хвостовой части отсутствуют реберные кости.
Между телами позвонков располагаются прослойки студенистой массы - остатки хорды, которые обеспечивают эластичность и упругость позвоночного столба. Таким образом, позвоночник не представляет собой единой кости. Он имеет вид цепочки, состоящей из жестких элементов - позвонков и эластичных дисков. Позвонки соединены между собой подвижно при помощи эластических связок. Такая конструкция позвоночного столба обеспечивает большую подвижность и упругость позвоночника в горизонтальной плоскости. Для рыб это очень важно, так как поступательное движение рыб достигается благодаря S-образным изгибам туловища и хвостового стебля.
Скелет головы имеет сложное строение и объединяет более 50 в основном парных костей (рис. 5.3). Он включает в себя кости черепа и висцеральную часть головы (кости верхней и нижней челюстей, 5 пар жаберных дуг и 4 кости жаберных крышек).
Периферический скелет представляют кости непарных плавников, кости поясов парных плавников, а также мускульные косточки. Основу непарных спинного и анального плавников составляют радиалии, к которым крепятся лучи плавников.
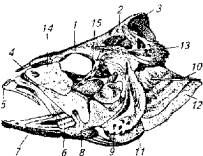
Рис. 5.3. Основные кости головы окуня:
1 - лобная; 2- теменная; 3- верхнезатылочная; 4- носовая; 5 - предчелюстная; 6 - верхнече-люстная; 7- зубная; 8- суставная; 9 - предкрышка; 10- крышка; 11 - межкрышка- 12- подкрышка; 13- задневисочная; 14- предглазничная; 15- глазничные кости
Парные плавники (рис. 5.4) - грудные и брюшные - имеют собственный скелет, который представлен костями свободного плавника и костями соответствующего пояса (плечевого или тазового). Плечевой пояс костистых рыб состоит из лопатки, коракоида, трех костей клейтрума и задневисочной кости. Задневисочная кость является элементом черепа и поэтому придает плечевому поясу прочность и относительную неподвижность, которая усиливается неподвижным соединением клейтрумов правой и левой половин тела.
Тазовый пояс (пояс брюшных плавников) с осевым скелетом жестко не связан. Он состоит из двух (правой и левой) треугольных костей, к которым крепятся плавники.
Костная основа грудных и брюшных плавников неодинакова. В состав грудных плавников входит три типа костных образований: базалии. множественные радиалии и плавниковые лучи.
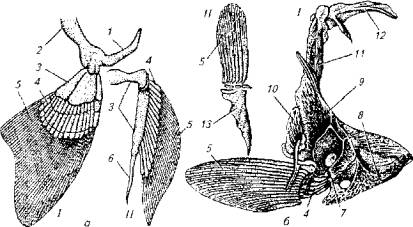
Рис. 5.4. Кости парных плавников и их поясов:
а-хрящевая рыба- б-костистая рыба; I-грудной плавнике плечевым поясом; II -брюшной плавник с тазовым поясом;1 - лопаточный отдел; 2- коракоидный отдел; 3-базалии; 4-радиалии; 5 -лучи плавников; 6- птеригоподии; 7-лопатка; 8- коракоид; 9-клерум; 10-задний клейтрум; 11 -надклейтрум; 12-задневисочная кость; 13-тазовая кость
В брюшных плавниках костистых рыб радиалии, как правило, отсутствуют. Следует подчеркнуть, что в целом опорная часть грудных плавников более совершенна. Они имеют и более развитую мышечную систему. Именно поэтому грудные плавники обеспечивают сложные поведенческие акты, о чем будет сказано отдельно
§21. МЫШЦЫ РЫБ
В организме рыб имеется три типа мышечной ткани: скелетная, гладкая и сердечная. Помимо топографии они различаются морфофункциональными свойствами. В этой главе обсуждаются физиологические особенности первых двух типов мышечной ткани рыб. Свойства сердечной мышцы рассматриваются в главе, посвященной кровообращению.
Скелетные мышцы обеспечивают сложные локомоторные реакции рыб в воде: поиск и потребление пиши, оборона или нападение, ритуальные движения во время нереста, миграции рыб и т. д.
Существенное влияние на характер движений рыбы оказывает форма ее тела (см. рис. 5.1). Она отражает этолого-экологические особенности рыб. Пелагические быстрые рыбы (скумбрия, тунец) имеют торпедообразное тело. У донных рыб (камбала, скаты) чаще всего уплощенное тело. Удлиненное тело облегчает передвижение по грунту или даже по суше (угорь) или обеспечивает проникновение в расщелины между камнями или норы (вьюн, мурена).
Перемещение в пространстве рыбы осуществляют за счет сокращения или туловищной мускулатуры (рис. 5.5), или мышц грудных плавников (рис. 5.6).
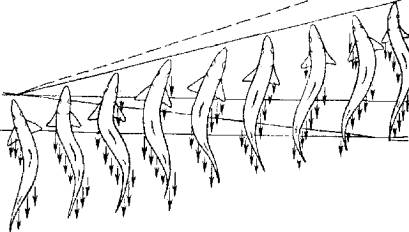
Рис. 5.5. Локомоторная волна туловища и хвостового стебля
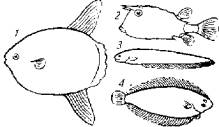
Рис. 5.6. Примеры движения рыбы при помощи мышц плавников:
1 - луна-рыба; 2- кузовок; 3 - электрический угорь; 4 -камбала
В первом случае рыбы отталкиваются туловищем, хвостовым стеблем и хвостовым плавником от воды. В результате изгибания и разгибания туловища в горизонтальной
плоскости возникает локомоторная волна. Такой механизм внешней кинематики лежит в основе движения большинства активных рыб. Этот тип движения имеет две разновидности: угревидную и скумбриевидную.
У рыб с удлиненным телом (угорь, вьюн, мурена) количество изгибов тела может быть больше одного. У рыб с вытянутым телом длина локомоторной волны все время остается постоянной: амплитуда колебаний тела от головы к хвосту не возрастает. Угревидный тип движения обеспечивает невысокие скорости перемещения, однако дает преимущества в энергетике, поскольку является более экономичным.
Известно, что самые быстрые пловцы (тунцы), развивающие скорость свыше 100 км/ч, имеют укороченное торпедообразное тело с хорошо дифференцированным хвостовым плавником. Такая форма тела обеспечивает рыбе минимальное гидродинамическое сопротивление, что существенно при высоких скоростях движения. Особо следует подчеркнуть роль хвостового плавника при скумбриевидном движении. Примерно 40 % мощности обеспечивают хвостовой стебель и плавник. Высокий и узкий хвостовой плавник тунцов и скумбрий способствует развитию больших скоростей плавания. Локомоторная волна от головы к хвостовому плавнику возрастает.
Кроме того, хвостовой плавник обеспечивает высокую маневренность рыбе на большой скорости. Так, карась при помощи хвостового плавника легко совершает поворот на 90њ за одно движение хвоста. При удалении хвостового плавника для выполнения этого же маневра рыбе требуется уже несколько взмахов хвоста. У многих рыб хвостовой плавник создает определенную подъемную силу, что важно при их отрицательной плавучести.
§22. МЕТАМЕРИЯ СКЕЛЕТНОЙ МУСКУЛАТУРЫ
Расположение скелетной мускулатуры у рыб носит сегментарный характер. Количество мышечных сегментов соответствует числу позвонков и колеблется (в зависимости от видовой принадлежности) от 25 до 100 сегментов и более. Отдельный мышечный сегмент у рыб принято называть миомером. В пределах одного миомера мышечные волокна расположены в краниально-каудальном направлении. Два соседних миомера разделяет миосепт - соединительнотканный сегмент, закрепленный на скелете рыбы.
Отдельное мышечное волокно в составе миомера прикрепляется к костям скелета при помощи миосептов, Миосепты у рыб выполняют функцию, аналогичную мышечным сухожилиям у высших позвоночных животных. Следует отметить, что скелетная мускулатура рыб лишена проприорецепторов - мышечных веретен. Однако соединительнотканные элементы мышц, включая миосепты, имеют нервные окончания, играющие роль тензорецепторов. Таким образом, обратная связь локомоторных центров с мышцами осуществляется посредством миосептальных проприорецепторов, степень возбуждения которых прямо пропорциональна степени возбуждения (сокращения) скелетных мышц.
С другой стороны, миосепты можно рассматривать как соединительные мостики между мышечными волокнами соседних миомеров. В единичный локомоторный акт вовлекается одновременно несколько соседних миомеров, что создает достаточное усилие для изгибания позвоночного столба. Чем больше миомеров возбуждается, тем больший изгиб совершает позвоночный столб и тем большее толчковое усилие создает хвостовой стебель.
Объемная геометрия отдельного миомера костистых рыб имеет довольно замысловатый вид. Только у бесчерепных рыб миомер можно рассматривать как более или менее правильный сегмент тела поперечной ориентации (рис. 5.7).
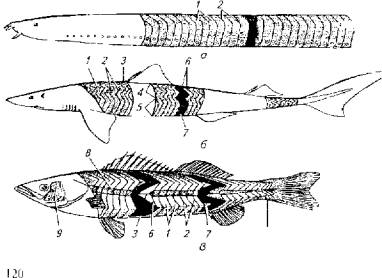
Рис. 5.7. Метамерия тела рыб:
а- круглоротые; б - пластинчатожаберные; в -высшие костистые рыбы; I и 2 - миомеры; 3- миосепты; 4 - спинная мускулатура; 5- латеральная мускулатура; 6 и 7- мышечные конусы; 8 и 9-красные мышцы; 10- мускулатура хвостового плавника
У пластинчатожаберных рыб миомер уже имеет зигзагообразную форму, причем зигзаги формируют большие тупые углы, У костистых рыб зигзаги миомеров образуют острые углы, отчего на поперечном срезе наблюдается рассечение сразу нескольких миомеров. Острые углы зигзагов миомеров формируют так называемые мышечные конусы, в состав которых попадают мышечные волокна (слои) от 4-6 соседних миомеров. Усложнение геометрии миомера, очевидно, обеспечивает преимущества в скорости и выносливости. Самая сложная метамерия тела обнаружена у активных пловцов - тунца и акул, у которых количество мышечных слоев в мышечных конусах достигает 20.
Скелетная мускулатура рыб морфологически и функционально разнородна. В ней различают красные (темные) и белые (светлые) мышцы (табл. 5.3).
Дата добавления: 2014-12-22; просмотров: 2345;