Абсолютная и относительная масса головного мозга и глаз у некоторых видов рыб (М. Ф. Никитенко, 1969)
| Вид рыб | Масса тела, г | Головной мозг | Глаза | ||
| масса, г | % от массы тела | масса, г | % от массь мозга | ||
| Лосось | 2,14 | 0,04 | 8,14 | ||
| Щука | 0,46 | 0,13 | 4,92 | ||
| Жерех | 0,62 | 0,04 | 3,91 | ||
| Сом | 2,83 | 0,05 | 3,83 | ||
| Налим | 0,96 | 0,08 | 1,62 | ||
| Угорь речной | 0,36 | 0,03 | 0,42 | ||
| Форель ручьевая | 0,18 | 0,21 | 1,01 | ||
| Окунь речной | 0,25 | 0,15 | 0,93 | ||
| Плотва | 0,21 | 0,26 | 1,02 | ||
| Карась | 0,40 | 0,60 | 0,92 | ||
| Сазан | 1,28 | 0,12 | 2,87 | ||
| Карп | 0,37 | 0,05 | 1,63 | ||
| Голавль | 0,16 | 0,25 | 0,88 |
Исключительно важная роль принадлежит ядрам боковой линии и вестибулярного аппарата. Поэтому продолговатый мозг рассматривают как коллектор акустико-латеральной афферентации. Помимо того, что они регулируют сложные двигательные акты, на этих ядрах замыкаются многие условные рефлексы рыб.
Вышеупомянутые гигантские маутнеровские клетки продолговатого мозга имеют большое количество синапсов, образованных с аксонами чувствительных нейронов (рис. 3.7). Эти клетки способны к особого рода интегральным процессам, нехарактерным для обычных нервных клеток.
Как следует из рис. 3.7, маутнеровская клетка получает информацию от аппарата акустико-латеральной системы (1 и 2). Эти клетки суммируют афферентацию и от других участков центральной нервной системы -- спинного мозга, мозжечка и среднего мозга. Таким образом, гигантские клетки продолговатого мозга являются своеобразными интегрирующими станциями.
Аксоны маутнеровских клеток уходят к передним мотонейронам и мотонейронам спинного мозга и осуществляют реципрокное торможение аналогичных мышц правой и левой сторон. Это позволяет рыбе производить удар хвостом или движения плавником при перемещении в продольном или вертикальном направлении (рис. 3.8).
Гигантские аксоны маутнеровских клеток образуют электрические синапсы с мотонейронами, ско-рость проведения возбуждения через которые очень высока (около 100м/с).
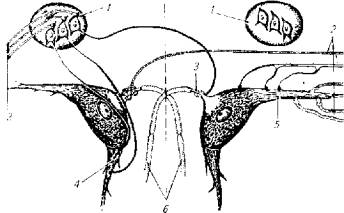
Рис.3.7. Гигантские маутнеровские клетки (5) продолговатого мозга и их связи с чувствительными нейронами (2, 3 и 4) и мотонейронами (6 - аксон, направленный на мотонейрон)
Синаптическая задержка при этом не превышает 0,05 м/с. Благодаря такой физиологической особенности гигантских клеток продолговатого мозга рыба совершает резкие броски. Поэтому значение маутнеровских клеток в поведении рыб заключается в обеспечении реакций тревоги и избегания большой опасности.
В продолговатом мозге находятся центры регуляции вегетативных функций - дыхания, пищеварения, кровообращения. При одностороннем разрушении продолговатого мозга наблюдается одностороннее прекращение дыхательных движений жаберных крышек.
В экспериментальных условиях за счет искусственной электрической стимуляции исследователи получают усиление или угнетение сердечной деятельности и Дыхания. Электрическая активность дыхательного центра продолговатого мозга изменяется в зависимости от газового состава воды, протекающей через жаберный аппарат.
При недостатке кислорода частота электрической ритмической активности дыхательного повышается. На этом фоне увеличение концентрации Углекислоты (диоксида углерода) в воде (и соответственно в крови) тормозит дыхательный центр. Повышение концентрации углекислоты (диоксида углерода) при нормальной обеспеченности кислородом не влияет на дыхательный ритм рыб.
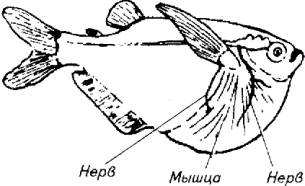
Рис. 3.8. Иннервация мышц грудного плавника у клинобрюшки с участием маутнеровских клеток
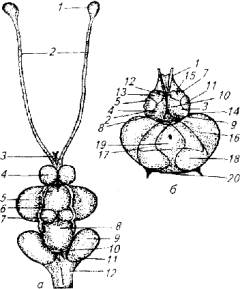
Рис. 3.9. Головной мозг карпа:
(а): 1 - обонятельная луковица; 2 - обонятельный тракт, 3-хиазма; 4-передний мозг; 5, 6- средний мозг; 7, 8- мозжечок; 9 - вагусная доля; 10,11 - лицевые доли; 12~- продолговатый мозг (б): 1 - обонятельный тракт; 2- 8 - борозды переднего мозга; 9-15-бугорки переднего мозга: 16, 17- средний мозг 18 - 20- мозжечок
У костистых рыб в составе продолговатого мозга выделяются ядра лицевого нерва (VII пара), что объясняется сильным развитием вкусового анализатора и органов общехимической рецепции. У сазана, карпа, пескаря, вьюна, сома именно они обеспечивают успешный поиск пищи. У остальных рыб афферентация от вкусовых почек поступает в продолговатый мозг по волокнам, входящим в состав блуждающего нерва.
Особое значение для регуляции вегетативных функций имеют ядра блуждающего нерва. У большинства карповых рыб на продолговатом мозге заметны крупные вздутия - вагусные доли (рис. 3.9), с базальной поверхности которых отходят многочисленные пучки блуждающего нерва. Крупный размер вагусных долей и ядер (особенно у карповых) подчеркивает исключительность X пары нервов в регуляции фи-зиологических функций рыб.
Блуждающий нерв, который делится на две ветви (боковой и жаберно-кишечный нерв), иннервирует все висцеральные органы как головной, так и туловищной части тела. Раздражение вагусных долей приводит к изменению функций органов дыхания, пищеварения и кровообращения.
Клетки, образующие ретикулярную формацию продолговатого мозга, обладают ритмической электрической активностью частотой 30 Гц. Однако ее частота может изменяться под влиянием афферентных потоков от экстеро-, проприо- и интерорецепторов и фактически всех сенсорных систем.
Поэтому изменения внешних условий отражаются на состоянии электрической активности ретикулярной формации, которая, в свою очередь, задает ритм дыханию, сердечным сокращениям, влияет на ритмическую активность спинного мозга. Большое влияние на физиологическую активность спинного мозга оказывает так называемая олива продолговатого мозга.
О функциональном значении этого образования свидетельствуют мощные проводящие пути, соединяющие оливу со спинным мозгом (спинооливный путь), с мозжечком (оливоцеребеллярный ПУТЬ), с промежуточным мозгом (таламооливный путь). Олива лучше развита у подвижных видов рыб с хорошо развитой мускулатурой хвоста и туловища. У высокоактивных пелагических рыб имеется добавочное ядро оливы.
Таким образом, продолговатый мозг у рыб выполняет роль интегрирующего отдела центральной нервной системы, обеспечивающего адекватную реакцию организма на изменения внешней среды, которая проявляется как в локомоторных, так и в вегетативных ответах. Особо надо отметить роль продолговатого мозга в выработке условных рефлексов у рыб.
Средний мозг. Появление у рыб хорошо развитого зрительного анализатора привело к формированию в головном мозге оптических путей, концентрирующих афферентный поток в среднем мозге (рис. 3.10).
У более организованных животных оптические пути' ветвятся и расходятся в область таламуса (зрительные бугры) и коры больших полушарий (затылочная область), оставляя за средним мозгом менее значимые функции, связанные с ориентацией глаза, адаптацией к изменению уровня освещенности.
У рыб все, что связано с функцией зрения, ассоциируется со зрительными долями среднего мозга. Даже процесс формирования временных связей при выработке условных рефлексов является прерогативой среднего мозга.
У костистых рыб в среднем мозге формируется примитивная кора (тектум) и тегментум. В ней происходит афферентный синтез. Сюда поступает афферентный поток не только от зрительного анализатора. Он имеет афферентные сообщения с продолговатым, промежуточным мозгом и мозжечком. К ядрам среднего мозга подходят афферентные пути от органов акустико-латеральной системы, органов обоняния, вкусовых почек, тактильных рецепторов органов общей химической рецепции, т.е. в среднем мозге собирается и суммируется афферентация со всех сенсорных систем рыб.
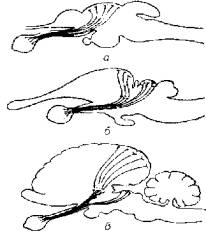
Рис. 3.10. Связи оптического пути со средним мозгом у рыб (а), рептилий (б) и млекопитающих (в)
В свою очередь, кора среднего мозга образует нисходящие пути к продолговатому мозгу (текто-бульбарный) и спинному мозгу (тектоспинальный). Мощный нервный пучок связывает средний мозг костистых рыб и с мозжечком (мезэнцефально-мозжечковый тракт). Эфферентная связь соединяет средний мозг с таламусом и гипоталамусом.
В коре среднего мозга высших рыб, несмотря на ее (в целом) примитивное строение, насчитывают до 10 слоев. Она (тектум) имеет проекции некоторых органов, что доказано при помощи электрической стимуляции ее различных участков.
В тегментуме среднего мозга находятся ядра III и IV черепно-мозговых нервов. К нему подходят также чувствительные волокна V и VIII нервов.
Электрическая стимуляция среднего мозга провоцирует определенные локомоторные реакции у рыб (рис. 3.11).
В среднем мозге рыб еще нет красного ядра, но тем не менее этот участок головного мозга имеет прямое отношение к поддержанию мышечного тонуса, о чем свидетельствуют опыты с перерезкой головного мозга выше и ниже среднего мозга.
Одностороннее разрушение среднего мозга вызывает нарушения реципрокной иннервации мышц туловища. При этом наблюдается сокращение мышц тела на стороне разрушения мозга, и рыба движется по кругу. Следовательно, средний мозг рыб отвечает за перераспределение тонуса антагонистических мышц - аналогичных мышц правой и левой сторон тела.
У спинальных рыб отмечаются спонтанные движения плавников, которые отсутствуют у рыб с интактным продолговатым и средним мозгом. Это свидетельствует о тормозящем влиянии среднего мозга на двигательные центры продолговатого и спинного мозга.
В среднем мозге замыкаются связи безусловной хватательной реакции со зрительной и акустико-латеральной афферентацией. Они определяют технику броска и захвата жертвы хищником (рис, 3.12).
В этой реакции задействованы мышцы тела, плавников и глаз. Возбуждение центра захвата формируется под влиянием сенсорной афферентации от органов боковой линии, слуха и зрения. Однако бросок осуществляется только после ориентации тела по отношению к жертве и попадания образа жертвы в область наибольшей остроты
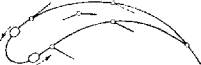
Рис. 3.11. Локомоторные ответы форели на электрическую стимуляцию разных участков среднего мозга (движения глаз,
плавников, хвоста)
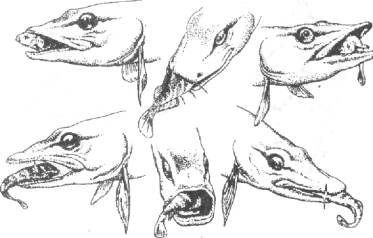
Рис. 3.12. Реакция захвата жертвы хищником
зрения на сетчатке глаза. Последнему способствуют не только мышцы тела (хищник занимает наиболее удобное положение для броска), но и движения глаз (возбуждение ядра глазодвигательного нерва среднего мозга).
Эволюционисты считают усложнение структуры среднего мозга и формирование его проводящих путей важным филогенетическим событием, поскольку оно стимулировало образование новых отделов головного мозга: промежуточного, полушарий переднего мозга.
Развитие среднего мозга послужило причиной качественного изменения функций мозжечка.
Мозжечок. У костистых рыб мозжечок хорошо развит (рис. 3.13), однако степень его развития зависит от этологических особенностей вида или популяции. У активных пелагических рыб, а также хищников он более развит по сравнению с малоподвижными, например донными, рыбами.
Мозжечок не только обеспечивает центральную регуляцию локомоторных реакций, но и наравне со средним мозгом берет на себя функцию интеграции деятельности нервных структур, Выше отмечались его связи со средним мозгом. Но помимо этого мощные нервные пучки соединяют мозжечок с продолговатым мозгом мозжечково-бульбарный путь) и лабиринтом (мозжечково-вестибулярный путь). В мозжечок тянут-ся волокна от ядер VIII и X нервов.
Удаление мозжечка может иметь неодинаковые последствия разных видов рыб. Так, у подвижных рыб (окунь, судак, щука) удаление мозжечка приводит к очень сильным изменениям. Помимо нарушения координации движений, тонуса мышц у этих рыб исчезают тактильная чувствительность, болевые ощущения, нарушаются зрение и слух. У донных малоподвижных рыб последствия удаления менее заметны.
Интересно, что у рыб, лишенных мозжечка или отдельных его частей (тела, заслонки), нарушается условно-рефлекторная деятельность. В этом случае условные рефлексы на световые и звуковые стимулы вырабатываются при значительно большем числе сочетаний. К тому же сохраняются такие рефлексы в тече-ние очень короткого времени. Установлено, что при удалении тела мозжечка исчезают ранее выработанные рефлексы. Таким образом, мозжечок следует рассматривать как орган, в котором происходит замыкание условных рефлексов с центрами зрительных органов и органов акустико-латеральной системы.
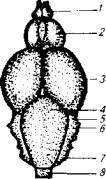
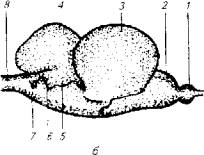
Рис. 3.13. Головной мозг лосося:
а-вид сверху; б-вид сбоку; 1 - обонятельная луковица; 2- передний мозг; 3- средний мозг; 4-мозжечок; 5-eminentia granularies; 6- n. trigeminus; 7-блуждающий нерв; 8-спинной мозг
Гибель безмозжечковых рыб наступает в результате трофических нарушений: изъязвления кожи, выпадения чешуи, нарушения васкуляризации, астении. Следовательно, мозжечок рыб имеет отношение и к регуляции вегетативных функций.
Промежуточный мозг. Этот мозг у рыб еще не дифференцировался в самостоятельную структуру в том виде, в каком он присутствует у высших позвоночных. Филогенетическое формирование промежуточного мозга тесно связано со средним и передним мозгом.
Развитый промежуточный мозг состоит их трех основных частей: дорзальной (эпиталамус, эпифиз), средней (таламус, или зрительные бугры) и вентральной (подбугорье, или гипоталамус).
Наиболее древней частью промежуточного мозга является дорзальный таламус - эпифиз. Он имеется у всех позвоночных, включая бесчерепных. Его изначальная функция сводилась к светорецепции. У костистых рыб роль эпифиза усложняется. Он превращается в орган чувствительной дифференциации. Электрическая активность эпифиза изменяется под влиянием зрительной, химической и электромагнитной стимуляции. При этом у рыб за ним сохраняется функция фоторецепции и эндокринного органа. У гольяна, например, местное освещение эпифиза приводит к потемнению тела за счет экспансии пигмента в хроматофорах.
К эпиталамусу можно отнести и еще одно древнее образование головного мозга - габенулярный узел. Он представлен парой ганглиев, связанных между собой комиссурой. Габенулярный узел имеет нервные связи как с передним мозгом, так и с эпифизом. Функционально он рассматривается как одно из центральных звеньев афферентного синтеза химической и зрительной информации.
Эволюционное развитие габенулярного узла тесно связано с выделением из состава среднего мозга его базальной части - таламуса. У большинства рыб он морфологически все еще зависит от среднего мозга. Его морфологическое обособление в зрительные бугры вызвано изменением образа жизни - развитием конечностей, выходом на сушу, повышением роли зрения. Поэтому зрительные бугры как самостоятельные морфологические структуры можно увидеть у животных, начиная с амфибий. Среди рыб только двоякодышащие с их хорошо развитым зрительным анализатором и примитивными конечностями имеют вздутия таламуса - зрительные бугры.
Связь таламуса с функцией движения отмечают и у рыб. По крайней мере, при его разрушении у рыб наблюдаются серьезные нарушения координации движений. Морфологической основой функциональной самостоятельности таламуса являются его дорзальные и вентральные ядра.
Афферентный поток поступает в таламус по обонятельным путям, а также нервным волокнам вкусовых и общехимических рецепторов. Таламус рыб осуществляет анализ и синтез афферентной информации, ее интеграцию, а также регулирует чувствительность организма.
Поскольку у рыб отсутствуют таламические зрительные бугры, То нет и подбугорья. Несмотря на то что гипоталамус у рыб еще не оформлен в самостоятельную морфологическую структуру и является вентральной частью среднего мозга, в функциональном отношении он самостоятелен. Его функциональную значимость подчеркивает множество афферентных и эфферентных связей, которыми гипоталамус соединен с передним, средним мозгом и таламусом. В гипоталамусе сходятся афферентные потоки от обонятельных, вкусовых, химических рецепторов, а также от органов акустико-латеральной системы.
В состав гипоталамуса входят особые нервные клетки, осуществляющие функцию рецепции. Экспериментально доказана чувствительность гипоталамуса к изменениям осмотического давления крови, рН, газового состава крови. Известна его реактивность к электромагнитным полям.
Сосудистый мешок гипоталамуса выстлан мерцательным эпителием, ворсинки которого чувствительны к продольным перемещениям рыбы. К гипоталамусу подходит терминальный нерв, функциональное предназначение которого неясно. Он берет своё начало в обонятельном эпителии носовых мешков и проходит параллельно обонятельному нерву. Известно мнение, что этот нерв отвечает за передачу информации от органов обшей химической рецепции в соответствующее (терминальное) ядро гипоталамуса.
Особое положение в составе промежуточного мозга занимает мозговой придаток - гипофиз, Гипоталамо-гипофизарный тракт объединяет придаток и промежуточный мозг в единую структуру, но с функциональными различиями. Гипофиз является нейросекреторным органом. Следует подчеркнуть, что это единственная железа внутренней секреции у рыб. Все остальные железы таковыми можно считать с большим допущением. Морфологически они не составляют систему железистых органов, так как представляют собой классический пример паракриновой секреции.
Таким образом, с одной стороны, гипоталамус можно рассматривать как коллектор афферентной информации, с другой - как центр интеграции ряда функций. Он связан афферентными и эфферентными путями с таламусом, средним мозгом, мозжечком и продолговатым мозгом. Эфферентные пути, идущие от гипоталамуса к ядрам продолговатого мозга (тройничного, слухового, блуждающего нервов), обеспечивают его регулирующую роль по отношению к висцеральным функциям. Поскольку у рыб большинство нервов содержат как парасимпатические, так и симпатические волокна, то становится понятным регулирующее влияние гипоталамуса рыб на вегетативные функции: работу сердца, дыхание, пищеварение и др.
В ядрах гипоталамуса обнаруживаются нейросекреторные клетки. Особенно высока секреторная активность трех ядер гипоталамуса: супраоптического, паравентрикулярного и ядра терминального нерва. Нейросекреция является общебиологическим явлением, так как характерна для животных разного уровня организации (рис. 3.14). Нейросекреты гипоталамуса (серотонин, эндорфины и др.) играют важную роль в процессе интеграции деятельности разных отделов нервной системы, формировании поведенческих мотиваций и запуске сложных инстинктов.
Нейросекреты гипоталамуса (релизинг-факторы) стимулируют синтез, накопление и секрецию тропных гормонов гипофиза {СТГ, ТТГ, АКТГ, ЛГ, ФСГ). В клетках гипоталамуса происходит синтез вазопрессина и окситоцина - гормонов, которые накапливаются в гипофизе. Тесные структурные и функциональные связи ядер гипоталамуса с гипофизом лежат в основе нейрогуморальной регуляции физиологических функций. Наиболее изучена эта связь в отношении функции размножения, развития адаптационного синдрома (стресс-реакция), течения ряда обменных процессов (прежде всего углеводного и жирового обмена). Гипоталамус посредством своих гуморальных агентов и нервных связей оказывает регулирующее влияние на все функции организма рыб.
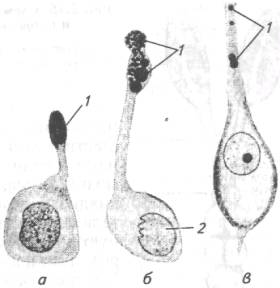
Рис. 3.14. Нейросекреторные клетки разных животных:
а- улитки; б-линя; в -человека; 1 -нейросекрет; 2-клеточное ядро
Промежуточный мозг у рыб играет существенную роль в афферентном синтезе ольфакторной и зрительной информации. Сюда же поступает информация от акустико-латеральной системы и тактильных рецепторов. Промежуточный мозг осуществляет дифференциацию разных видов чувствительности. Не исключено, что он причастен к анализу и афферентному синтезу электромагнитной рецепции, которая играет важную роль в формировании навигационных способностей мигрирующих рыб.
Промежуточный мозг имеет отношение к образованию условных рефлексов. Его повреждение отражается на скорости выработки рефлексов и проявлении ранее выработанных рефлексов. Однако механизм участия промежуточного мозга в выработке условных рефлексов неясен. Не исключено, что образование условного рефлекса сопряжено с нейросекреторной активностью мозга. Однако нельзя исключать и другое: промежуточный мозг просто повышает (а его разрушение понижает) возбудимость других отделов головного мозга к определенным условным раздражителям.
Передний мозг. У костистых рыб эта часть головного мозга представлена двумя группами структур (рис. 3.15). Это прежде всего обонятельный мозг, в состав которого входят мантия (плащ) и гиппокамп (первичная мантия). Однако довольно большая часть Среднего мозга рыб (пограничный участок между передним и промежуточным мозгом) не имеет отношения к ольфакторной рецепции. Это полосатые тела, которые составляют основную массу среднего мозга рыб.
Несмотря на то что полосатые тела рыб - структуры крупные, их физиологическая роль до конца неясна. Очевидно, что одной проводниковой функции для оправдания существования полосатых тел недостаточно. Однако среди афферентных связей переднего мозга описана только связь с обонятельным анализатором. У высших животных полосатые тела как часть базальных ганглиев играют важную роль в регуляции сложных поведенческих реакций, а в качестве составного элемента лимбической системы определяют эмоциональное состояние животного. Однако у одиночных костистых рыб удаление переднего мозга никак не отражается на их двигательной активности. Мантия и гиппокамп выполняют роль высшего обонятельного центра.
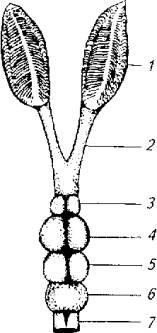
Рис. 3.15. Схема, иллюстрирующая соотношение переднего и головного мозга в целом у европейского угря:
1- обонятельная розетка; 2 - обонятельный нерв; 3- обонятельная луковица; 4- передний мозг; 5-средний мозг: 6- мозжечок; 7- продолговатый мозг
Мантия рассматривается физиологами как конечная станция вторичных обонятельных волокон, идущих от обонятельных луковиц. Эта часть ольфакторного анализатора хорошо развита у рыб, для которых обонятельная сенсорика выходит на первый план во взаимодействиях с внешней средой. Так, у европейского угря обонятельный мозг занимает значительную часть головного мозга. Абсолютный размер луковиц составляет 3-5мм, а масса- 10- 15мг (рис. 3.15).
Обонятельный мозг угря в функциональном отношении очень активен. Порог обонятельной чувствительности у европейского угря в 1000 раз превышает таковой гольяна и в 100 раз - форели. Столь высокая чувствительность угря к запахам позволяет исследователям считать передний мозг основным отделом центральной нервной системы, обеспечивающим навигационный поиск при миграциях рыбы.
Интересно, что целостность переднего мозга является обязательным условием для проявления группового поведения. После удаления переднего мозга у стайных рыб оперированные рыбы начинают плавать в одиночку. При этом нарушается и процесс стайного обучения, т.е. выработка условных рефлексов у группы рыб. У некоторых рыб (например, Lepomis cyanellus - американского ушастого окуня), обладающих высокой агрессивностью, разрушение переднего мозга приводит к превалированию миролюбивых стереотипов поведения, т.е. снижает агрессию.
Очевидно, что передний мозг рыб играет важную роль в формировании приспособительных реакций. Его развитые нервные связи с промежуточным и средним мозгом обеспечивают соответствующие влияния на высшие интегративные органы рыб. Передний мозг рыб является регулятором рефлекторных ответов за счет способности понижать порог чувствительности к разнообразным стимулам внешней среды. Таким образом, передний мозг создает предпосылки для развития процесса облегчения в ассоциативных отделах центральной нервной системы рыб.
Дата добавления: 2014-12-22; просмотров: 3454;