Химический состав магнетитных кристаллов тунца
Оксид Массовая доля, %
FeO 86,3
МnО 0.2
СаО 0,2
Всего 86,7
Магнетитные кристаллы находятся во взаимодействии друг с другом посредством собственных магнитных полей. При изменении внешнего магнитного поля отдельные кристаллы способны поворачивать-ся наподобие стрелки компаса, изменяя при этом свое собственное поле и суммарное поле решетчатой кости.
Ферромагнитная гипотеза магниторецепции позволяет объяснить реактивность рыб на магнитные поля и использование рыбами магнитных полей для навигации. Однако до сих пор не описана анатомическая структура, в которой происходит трансформация магнитного поля в потенциал действия, т.е. в нервный импульс. Гипотетически магниторецептор рыбы может иметь следующую схему (рис. 2.30).
Поворот кристалла магнетита раздражает чувствительное окончание дендрита нейрона. В результате образовавшийся потенциал действия возбуждает нейрон.
В магнитной решетке решетчатой кости ориентация и величина магнитного напряжения отдельных кристаллов магнетита генетически детерминированы. Однако экологические условия, в которых растет молодь, могут внести поправку в структуру решетки и напряженность кристаллов.
Суммарное магнитное напряжение магнитной решетки рыбы может быть достаточно высоким. Поэтому изменение напряженности магнитного поля рыбы, например, при изменении солнечной активности может привести ее в состояние тревоги, дискомфорта. Отсюда, снижение кормовой активности рыб, что рыбаки оценивают как отсутствие клева.
Магнитная решетка может выполнять и функцию своеобразной навигационной карты. Перед миграцией магнитное напряжение отдельных кристаллов магнетита и суммарное магнитное поле всей решетки настраиваются относительно магнитных линий Земли на пути предстоящей миграции. Отклонение от генетически детерминированного маршрута приводит к напряжению магнитного поля рыбы,
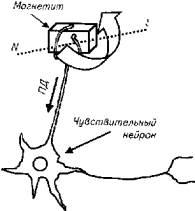
Рис.2.30. Гипотетическая схема магниторецептора
что она оценивает как дискомфортное состояние. Выйти из него можно лишь одним способом - привести кристаллы решетки в исходное напряжение, а это, в свою очередь, возможно только через изменение положения тела относительно магнитных линий Земли, т.е. рыба вынуждена вернуться на заданный маршрут.
Наличие магниторецепторов решетчатой кости объясняет реактивность к электромагнитным полям неэлектрических и слабоэлектрических видов рыб.
У сильноэлектрических видов рыб рецепция магнитного поля осуществляется боковой линией рыб и производными от нее структурами. В магнитном поле тело рыбы является источником индукционных электрических полей, которые фиксируются структурами боковой линии. В опытах на скатах показано, что электрическая активность ампул Лоренцини изменяется как в электромагнитном поле, так и в поле постоянного магнита.
Интересно, что реакции рыб на изменение магнитного поля зависят еще и от движения воды. Так, у ската реакция на магнитное поле в искусственном водоеме возникала тогда, когда ампулярный канал рецептора (ампулы Лоренцини) находился под углом к направлению тока воды. Если канал располагался вдоль водного потока, электроактивность ампул Лоренцини на изменение магнитного поля не регистрировалась. Следовательно, морские течения при миграциях рыб могут выполнять функцию корректировки направления движения рыб.
Некоторые специалисты высказывают мнение о том, что кроме описанных выше структур лабиринт является морфологической основой вероятной магниторецепции. Однако экспериментальных доказательств участия полукружных каналов в магниторецепции у рыб для этого недостаточно. Их связь с рецепцией магнитных полей у диких перелетных птиц и почтовых голубей убедительно доказана многочисленными экспериментами. Имеются указания и на то, что изменение напряженности магнитного поля приводит к изменению возбудимости узлов симпатической нервной системы без промежуточной магниторецепции.
Известно, что магнитное поле влияет на движение любого электрического заряда или частицы. Следовательно, реакция на магнитное поле организмом осуществляется и без специфических рецепторов. Мембранный потенциал, круговые токи, электрические явления в сердечной мышце и в нейронах изменяются в магнитном поле. Электрочувствительные органы также могут информировать об изменении магнитного поля. Изменения самочувствия человека, поведения домашних животных при изменениях геомагнитной обстановки общеизвестны. Изменение электромагнитного поля Земли перед глобальными катастрофами - землетрясениями, извержениями вулканов, ураганами- сопровождается этологическими аномалиями животных разного уровня организации (от муравьев до приматов). Массовую гибель животных, как и появление новых видов на Земле, многие исследователи связывают именное внезапно возникшими электромагнитными аномалиями, которые лишают животных пространственной и временной ориентации.
Магнитная афферентация, как и всякая другая сенсорная информация, поступает в промежуточный мозг. Вероятно, к магнитосенсорике имеет отношение эпифиз. У почтовых голубей, морских свинок и крыс наблюдали повышение электрической активности эпифиза в искусственном магнитном поле. У крыс искусственное магнитное поле изменяло секреторную активность эпифиза. В ночное время 15-минутная магнитная экспозиция повышала активность фермента ацетилтрансферазы и образование гормона мелатонина в эпифизе. Таким образом, таламус получает информацию об изменении геомагнитного поля по двум традиционным каналам - нервному и гуморальному.
Принимая во внимание, что электроактивность эпифиза возрастает и при световой стимуляции, можно предположить, что эпифиз причастен к афферентному синтезу при позиционировании рыбы в процессе навигации. При этом магнитосенсорная афферентация может играть ключевую роль.
Таким образом, теллурические токи, магнитные поля и флуктуация электромагнитного поля Земли, морские течения, световые и химические раздражители, а также соответствующие им сенсорные органы создают объективные предпосылки для механизма точного географического позиционирования и навигации у мигрирующих рыб.
§9. ТЕРМОРЕЦЕПЦИЯ
Температура воды для рыб как пойкилотермных животных имеет исключительное значение. У рыб активность ферментных систем проявляется в широком диапазоне температур. Поэтому и жизненная активность рыб не прекращается в осенне-зимний период, когда температура их тела достигает 4 њС, а у полярных - более низкой отметки.
Температурный диапазон природных водоемов очень широк, причем в большинстве водоемов присутствует ихтиофауна. Так, рыба семейства харацинидовых Cyprinodon macularis обитает в горячих источниках Калифорнии, выбирая участки водоема с температурой 52 њС, в то время как антарктическая сайка активна при отрицательной температуре воды (минус 2 њС). Известно, что обычный карась в нашей полосе вмерзает в лед, а после размораживания оживает, т. е. ихтиофауна приспособилась к самым разнообразным температурным условиям. Тем не менее, ихтиологам известны случаи массовой гибели рыб, которые не смогли приспособиться к изменениям температуры среды обитания. Причем гибель возможна как при понижении, так и при повышении температуры воды. В литературе описан случай массовой гибели рыбы Lophalotilus chamaeleonticeps у берегов Новой Англии в 1882 г. в результате понижения температуры прибрежных вод. Этот вид рыб стал вновь появляться у берегов Новой Англии лишь спустя 70 лет.
Вместе с тем гибель холодноводных рыб (форель, белорыбица) при повышении температуры воды не является редкостью.
Рыб делят на стенотермных, приспособленных к узкой амплитуде колебаний температуры окружающей среды, и эвритермных, выдерживающих большие перепады температур. У рыб высоких широт обмен веществ не угасает в широком диапазоне температур. Например, карп при температуре 1 њС обходится минимальной концентрацией кислорода в воде 0,8 мг/л, а при температуре 30 њС - 1,8 мг/л. Однако у таких видов, как сиги, таймень, налим, повышение температуры на 7-10 њС снижает активность обмена веществ.
Естественно, что все виды рыб имеют зону температурного комфорта и предпочтительные температуры. Например, для тиляпии это 28-30 њС, для карася и карпа 25 њС, для ельца 20 њС, для радужной форели 15-18 њС, а для беломорской трески всего 9 њС.
Причем у многих пресноводных европейских видов имеется летний и зимний температурный пре-ферендум. Например, карп, плотва, лещ летом отдают предпочтение участкам водоема с температурой 25-27 њС, а в осенне-зимний период - 2-4 њС.
Питаются рыбы при узком температурном интервале, который ограничен 10-15 њС. Наибольшая интенсивность питания у большинства рыб средних широт приходится на 10 - 22 њС. Рыбы высоких широт при повышении температуры воды испытывают дискомфорт; максимум их пищевой активности наблюдает-ся в довольно узком диапазоне температур - от минус 1 до плюс 4 њС.
Рост рыб также ограничен температурой среды. Температурный интервал определяется всего 5-7 њС. За пределами этого "ростового" коридора рыба проявляет пищевую активность при отсутствии роста.
С температурой среды связана и функция размножения (табл. 2.9). В зоне с резкими сезонными пе-репадами температур для большинства рыб сигналом к нересту является не только увеличение светового дня, но и весенний прогрев водоема. Исключения здесь редки (лососевые, налим).
Многие черноморские рыбы реагируют на повышение температуры воды даже на 0,03 "С. Карась реагирует на изменение температуры на 0,05 њС. Большинство видов рыб средней полосы проявляют меньшую термочувствительность, которая не превышает десятых долей градуса Цельсия.
Дата добавления: 2014-12-22; просмотров: 1631;