Нуклеиновая кислота→ Фермент → Продукт

Нуклеиновая кислота→ Фермент → Продукт
 |
Механизмы регуляции, замедляющие или ускоряющие реакции отдельных путей обмена веществ, связанны с переключением процесса обмена с одного пути на другой. Регулирующие факторы весьма разнообразны, включая образующиеся в обмене веществ промежуточные продукты, поступающие с пищей неорганические и органические вещества, также факторы внешней среды, как свет и температура.
Для многоклеточного организма системы регуляции разделяют на внутриклеточные, межклеточные и организменные.
К внутриклеточной системе относятся ферментная регуляция (на уровне ферментов), генетическая, мембранная и др. Межклеточные системы включают по меньшей мере гормональную, электрофизиологическую, трофическую.
На организменном уровне скорее всего следует говорить об организменном уровне интеграции.
Рассмотрим следующие основные типы клеточной регуляции: метаболитную, ферментную, генетическую, мембранную, гормональную, фоторегуляцию, электрическую (электрофизиологическая).
Метаболитная регуляция (регуляция путем изменения концентраций метаболитов, не затрагивающие активности или числа ферментных молекул).
Метаболитная регуляция основывается на двух основных принципах регуляции:
 |
а)
б)
а) Метаболит (х) в качестве отрицательного эффектора тормозит свой собственный синтез (по типу обратной связи).
б) Метаболит, выполняющий роль положительного эффектора, ускоряет свое собственное преобразование (по типу управления по возмущению).
Оба принципа имеют одинаковое назначение помешать образованию лишнего продукта.
При метаболитной регуляции на разветвлении путей обмена веществ выбор пути может определяться концентрацией общего для конкурирующих ферментов субстрата.
Высокие концентрации СО2 (как метаболита в широком смысле) стимулируют карбоксилирование, низкие – декарбоксилирование пировиноградной кислоты:
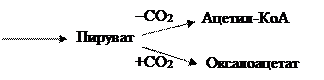 |
Поскольку ткани зеленых растений в течение дня испытывают недостаток СО2 (используется при фотосинтезе), а ночью обогащаются углекислотой (образуется при дыхании), то процессы карбоксилирования происходят главным образом ночью.
Наилучшим примером метаболической регуляции по принципу обратной связи может служить дыхательный контроль – зависимость интенсивности дыхания от концентрации АДФ.
 |
В результате энергопотребляющих реакций (различные синтезы, поглощение веществ, рост и т. д.) концентрация АТФ уменьшается, а АДФ соответственно возрастает (АТФ → АДФ + Фн). Посколько действие дыхательной цепи обязательно связано с фосфорилированием (АДФ + Фн → АТФ) и лимитируется количеством АДФ (дыхательное сопряжение), накопление АДФ при энергопотребляющих процессах вызывает ускорение действия дыхательной цепи и дыхательного процесса в целом и, таким образом, усиление генерации энергии в клетке. В этом случае мы сталкиваемся с исключительно экономичной формой регуляции количества доступной энергии путем «спроса».
Особое место в клетке занимают ферментные регуляции, поскольку все биохимические реакции носят ферментативный характер.
Ферментные регуляции (регуляции путем изменения активности имеющихся молекул ферментов), – регулирующие факторы в этом случае действуют непосредственно на фермент. Однако, все ферменты и соответственно этапы обмена веществ подчинены регуляторным механизмам. Наиболее эффективно регуляция выражена у ферментов, катализирующих ключевые реакции (ключевые ферменты), и в точках разветвления обменных процессов. Ключевые ферменты – это ферменты, катализирующие самые медленные реакции в какой либо цепи обмена веществ.
Ферментная регуляция затрагивает только один фермент, но происходит очень быстро (доли секунд) и служит для «тонкой настройки» путей обмена веществ.
Ферментная регуляция может осуществляться несколькими путями:
1. Обратимым или необратимым превращением неактивных предшественников в активные ферменты.
2. Изменением активности ферментов путем воздействия на его центры (изостерические и аллостерические эффекты, которые являются самыми важными).
3. Воздействием на процессы распада самих ферментов (например, ферменты могут попасть под атаку протеолитических ферментов).
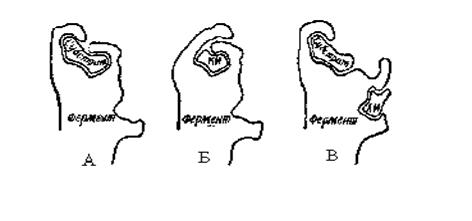
Большое значение для регуляции обмена веществ имеют изостерические и аллостерические эффекты, вызываемые взаимодействием фермента с малыми молекулами (эффекторами). Эффектор может действовать на молекулу фермента или связываясь с каталитическим центром и незначительно изменяя структуру фермента (изостерический эффект), или путем изменения третичной структуры всей ферментной молекулы (аллостерический эффект) (рис. 1.6).
|
| Рис. 1.6. Модель механизма изостерического (Б) и аллостерического (В) ингибирования: А – фермент в активном взаимодействии с субстратом. Б и В – фермент неактивен. КИ – конкурентный ингибитор. АИ – аллостерический ингибитор |
Изостерический эффект наблюдается в том случае, когда структуры молекул эффектора регулируемого фермента и его субстрата близки и поэтому эффектор может связываться с каталитическим центром, не претерпевая при этом никаких превращений. Таким образом, субстрат и эффектор конкурируют за молекулу фермента и такой тип ингибирования называют конкурентным ингибированием, причем эффектор является конкурентным ингибитором.
Будучи в избытке, конкурентный ингибитор вытесняет субстрат из мест связывания с ферментом; возникающее в результате торможение активности может быть снято большим количеством субстрата. Такой зависящий от концентрации веществ антагонизм отношения субстрат-ингибитор так же характерен для конкурентного ингибирования. Конформация ферментной молекулы при этом изменяется не больше, чем при связывании субстрата с ферментом.
Типичным примером конкурентного ингибирования является торможение сукцинатдегидрогеназы (субстрат НООС–СН2–СН2–СООН) малоновой кислотой (НООС–СН2–СООН), с помощью которой удается затормозить весь лимоннокислый цикл (цикл Кребса).
Аллостерические эффекты, основаны на функционировании ферментов, которые представляют собой полимерные белки, обычно состоящие из идентичных полипептидных субъединиц, с аллостерическими центрами. Активность таких ферментов регулируется не их субстратами, а другими веществами, присоединяющимися к ферментам в особых участках, удаленных от активного центра. Эти вещества влияют на активность фермента, вызывая обратимое изменение в структуре активного центра. Называются такие вещества аллостерическими эффекторами. В зависимости от характера влияния, которое они оказывают, увеличивая или уменьшая сродство фермента к субстрату, эффекторы подразделяются на аллостерические активаторы (ускоряющие реакцию) и аллостерические ингибиторы (тормозящие реакцию).
Аллостерические ферменты имеют два различных типа активных центров: регуляторный и каталитический.
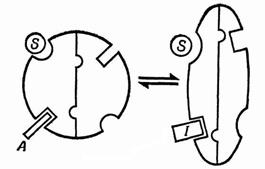
Схема, поясняющая работу аллостерического фермента приведена на рис. 1.7.
|
| Рис. 1.7. Механизм аллостерического эффекта: слева – активатор А стабилизирует более активное конформационное состояние фермента; справа – ингибитор I стабилизирует менее активное состояние; S – субстрат |
Примером аллостерического фермента может служить фосфофруктокиназа, катализирующая фосфорилирование фруктоза-6-фосфата с образованием фруктоза-1,6-дисфосфата. Эта реакция протекает во время гликолиза, составляющего одну из стадий процесса дыхания.
АТФ, если его концентрация высока, аллостерически ингибирует фосфофруктокиназу. Когда же клеточный метаболизм усиливается, а значит, расходуется АТФ и его общая концентрация падает, данный метаболический путь снова вступает в действие.
Когда конечный продукт какого-либо метаболического пути начинает накапливаться, он может действовать как аллостерический ингибитор на фермент, контролирующий первый этап этого пути. При этом снижается сродство данного фермента к его субстрату и соответственно уменьшается или вовсе приостанавливается дальнейшее образование самого конечного продукта. Это явление – ингибирование конечным продуктом представляет собой пример регуляции по принципу обратной связи.
Генная регуляция (регуляция путем включения или выключения синтеза ферментов) – регулирующие факторы действуют на генетический материал (ДНК) или непосредственный его продукт (РНК).
Генная регуляция, затрагивающая несколько ферментов одновременно, более экономична по сравнению с ферментной, поскольку ферменты, в которых нет надобности, просто не синтезируются. Генная регуляция занимает больше времени, так как включается трансляция и чаще всего также и транскрипция. Эта форма регуляции служит для «грубой настройки» обмена веществ и играет важную роль в процессах развития.
Физиологическая функция генов заключается в передаче информации клетке через мРНК и ферменты. Никогда не происходит одновременной передачи всей имеющейся информации, т. е. никогда не синтезируются одновременно все потенциально возможные ферменты. Таким образом, существуют одновременно активные (продуцирующие РНК) и неактивные гены. Благодаря генной регуляции осуществляется активация генов (индукция) и инактивация (репрессия); одним из самых первых, экспериментально уловимых, специфических последствий генной регуляции является появление или исчезновение того белка (фермента), синтез которого контролируется регулируемым геном (индукция и репрессия ферментов).
Для регуляции на уровне генов справедливы те же принципиальные закономерности, как и при ферментной регуляции. Активации ферментов субстратом соответствует генетическая субстратная индукция, торможению фермента конечным продуктом – репрессия генов продуктом реакции; оба механизма регуляции в равной мере запускаются метаболитами, различны лишь уровни, на которых осуществляется регуляция.
Экспериментально можно отличить генные регуляции от ферментных потому, что первые в отличие от вторых выключаются под действием ингибиторов синтеза белка и РНК (актиномицин, 6-метилпурин, пуромицин, этионин и др.).
Гормональная регуляция. Фитогормоны – соединения с помощью которых осуществляется взаимодействие клеток, органов, тканей и которые в малых количествах необходимы для регуляции физиологических и морфологических процессов. Гормоны – органические молекулы, которые образуются в тканях и органах и действуют в малых концентрациях – 10–11 моль/л и ниже. Образуясь в одной ткани, они перемещаются к другой и благодаря специфическим реакциям в ткани-мишени контролируют такие процессы как рост, развитие и дифференцировка. Выделяют пять основных классов фитогормонов: ауксин, гибберилины, цитокинины, АБК и этилен. Сегодня к фитогормонам относят еще брассиностероиды, жасминовую, салициловую кислоты, системин а также простагландины.
Мембранная регуляция. Мембранная регуляция очень хорошо прослеживается по функции мембран, которые мы рассмотрели на прошлой лекции. Отметим, что этот путь регуляции осуществляется, в основном, благодаря изменениям в мембранном транспорте. Так изменения в проницаемости мембран клеточных компартментов может стать фактором регуляции. Примером может служить сокращение митохондриальной мембраны при повышении концентрации АТФ (примета малой потребности в энергии). В результате мембрана становится менее проницаемой и тормозится приток продуктов гликолиза в митохондриях и соответственно дальнейший синтез АТФ.
Особое место в механизмах мембранной регуляции имеет система мембранных хема-, фото- и механорецепторов, позволяющих клетке отмечать изменения во внешней и внутренней среде и в соответствии с этим изменять свойства мембраны.
Но мембраны не только создают компартменты в клетке. Хорошо известно, что большинство клеточных ферментов встроены в мембраны, при этом каждый тип мембраны содержит свои ферменты, что обеспечивает ход соответствующих реакций. Более того, изменение проницаемости очень тесным образом связано с активностью мембранных ферментов.
Фоторегуляция. У растений существует белковая система, рецептирующая красный и дальний красный свет – фитохромная система. Фитохром присутствует в клетке в очень малых количествах, поэтому для запуска контролируемых им фотоморфогенетических реакций достаточна малая энергия. К таким реакциям относится: диэтиолирование проростков, выращенных в темноте, стимуляция прорастания некоторых семян, начало цветения и переход в состояние покоя.
Некоторые фотоморфогенетические реакции требуют значительно большей энергии света и вызываются преимущественно синим и далеким красным светом. Эти реакции могут регулироваться фитохромом либо пигментом, поглощающим синий свет – флаовопротеидом.
В природных условиях фитохром, возможно, контролирует форму растений и прорастание семян под пологом листьев, так как листья очень сильно поглощают свет в области 660 нм, но относительно слабо – при 730 нм. Состояние фитохрома может регулировать образование и количество фитогормонов четырех типов – этилена, цитокининов, ауксина и гиббереллина – в разных органах растений. Кроме того, длительное освещение иногда контролирует уровень абсцизовой кислоты. В некоторых случаях внесение гормона может снимать эффект трансформации фитохрома (например, действие гиббереллина при диэтилировании).
Остановимся еще на одной очень интересной системе регуляции – электрической или электрофизиологической. Как известно, у растений существует разность электрических потенциалов как между различными тканями, органами, так и между внутренней и наружной средой клетки, при этом внутренняя среда имеет отрицательный заряд по отношению к внешней. РЭП, например, между цитоплазмой и внешней средой составляет для растительной клетки порядка –150 – –300 мВ. Как правило, на каждой биологической мембране существует РЭП, которую называют мембранным потенциалом.
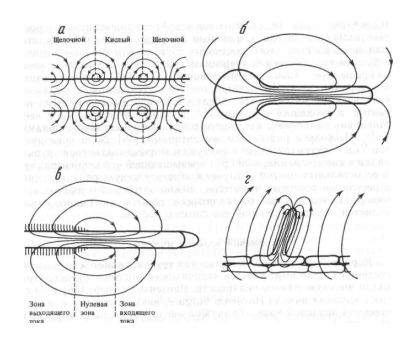
Необходимо отметить, что около растительных клеток и отдельных органов зарегистрированы электрические токи, которые в зависимости от рассматриваемого объекта имеют величины от 0,5 до 3,0 мкА см2. Эти токи зарегистрированы для одноклеточных харовых водорослей, прорастающих пыльцевых зерен (лилия), корневых волосках, колеоптилях, гипокотелях и т. д. (рис. 1.8).
|
| Рис. 1.8. Некоторые примеры околоклеточных (трансклеточных) электрических токов у растительных объектов: а – интернодальная клетка Chara с чередованием щелочных и кислых поясков; б – прорастающее пыльцевое зерно Lilium; в – первичный корень Hordeum; г – корневой волосок Hordeum. Стрелками показано направление движения положительных зарядов |
Во всех случаях биогенные электричные токи, окружающие и пронизывающие клетки, функционально очень тесно связаны с их ростом. Во всех растительных объектах околоклеточные или трансклеточные токи входят в растущую часть клетки и выходят с противоположной нерастущей. Это дало возможность называть эти токи «ростовыми». В целом корне они входят в зону роста (растяжением) и выходят из нерастущих частей ризадермальных клеток зоны диференцировки. В одном из опытов в трех из ста исследуемых волосков не были обнаружены околоклеточные токи. Оказалось, что у волосков отсутствует рост.
Но эти токи не просто связаны с ростом, они индуцируют его. Это очень хорошо показано на зиготе одного из видов бурой водоросли. При одностороннем освещении около зиготы регистрируются электрические токи, которые выходят из осветленного полюса и входят в затемненный. Таким образом, создается электрическое поле, по которому положительные заряды двигаются внутрь клетки снизу вверх, а отрицательно заряженные везикулы аппарата Гольджи (которые поставляют строительный материал для роста растяжением) – в противоположном направлении, сверху вниз. В результате в затененной нижней части возникает вырост, начинается рост растяжением. Таким образом, освещение индуцирует ток, а ток инициирует рост, создавая ось симметрии и тем самым задает вектор роста, его направление. Затем первое деление происходит поперек этой оси: нижняя дочерняя клетка дает начало ризоиду, верхняя –таллому (рис. 1.9).
|
| Рис. 1.9. Индукция односторонним освещением биогенных электрических токов и их связь с ростом и дифференциацией зиготы Pelvetia fastigiata: Темные кружки – протонные насосы; светлые прямоугольники – катионные каналы; замкнутые стрелки – силовые линии положительного тока; пунктирная линия – место первого деления |
Электрические токи не только индуцируют начало роста, они контролируют и происходящий рост. Основа этой регуляции – высокая чувствительность мембранных транспортных систем к изменению внешних и, как видно, внутренних условий.
Если принять толщину мембраны 10 нм (для плазмалеммы) она составляет 7,5 нм и величину мембранного потенциала ~ 100 то образуется очень высокая напряженность – ~ 500 В. Изменение такой величины напряженности не может не влиять на мембранные структуры, активность ферментов и т. д.
Известно также, что кроме электрических явлений в обычных условиях (в состоянии покоя) существуют, как и у животных, потенциалы действия, которые возникают при разных воздействиях на растительную клетку и организм в целом. Об этом мы упоминали и поговорим позднее при рассмотрении механизмов движения у растений.
Трофическая регуляция – воздействие с помощью питательных веществ – наипростейший способ связи между клетками, тканями и органами.
В корни и другие гетеротрофные органы растений поступают ассимиляты, образующиеся в листьях в процессе фотосинтеза. Надземные части, в свою очередь, нуждаются в минеральных веществах и воде, поступающих из почвы. Корни используют ассимиляты, поступающие из побега, на собственные нужды, а часть трансформированных органических веществ передвигается в обратном направлении. Таким образом, ассимиляты осуществляют связь между их поставщиками и потребителями.
Изменения в содержании разных элементов питания оказывают влияние на обмен веществ, физиологические и морфогенетические процессы в растениях. Хорошо известно, что изменения в растительном организме обусловлены также недостатком тех или иных минеральных элементов.
Дата добавления: 2014-12-22; просмотров: 1630;