Мозговая основа ощущений
Психологам с 20-х гг. известно, что ощущение возникает довольно поздно – после 100 мс от момента предъявления стимула (т.е. значительно позже прихода сенсорных импульсов в кору). В настоящей работе была поставлена задача понять, что происходит за это время и какой стадии мозговых процессов соответствует возникновение субъективного образа. В исследовании при предъявлении одних и тех же стимулов записывали объективные показатели мозговой деятельности в виде вызванных потенциалов и определяли количественные показатели восприятия. Для получения последних использовались методы теории обнаружения сигнала [Swets et al., 1961], которая описывает процесс восприятия с помощью двух независимых переменных: показателя сенсорной чувствительности d' и критерия решения, определяемого мотивационными факторами. Основные выводы построены на вычислении корреляций между показателями физиологии и психологии, а именно амплитудой волн вызванного потенциала и психофизическими индексами. Исследования проведены на соматосенсорном [Иваницкий, Стрелец, 1976] и на зрительном [Иваницкий, Матвеева, 1976] анализаторах, причем полученные на обеих модальностях результаты были принципиально сходными.
Было установлено, что амплитуда ранних волн вызванного потенциала (ВП) обнаруживала статистически достоверную корреляцию с показателем d', а поздних волн – с критерием решения. Промежуточные волны с латентностью 140 мс для соматосенсорного и 180 мс для зрительного анализатора коррелировали с обоими перцептивными индексами, причем такая двойная корреляция обнаруживалась только для волн проекционной коры. Амплитуда этих волн определялась, таким образом, как сенсорными свойствами стимула, так и его значимостью. На основании имеющихся данных о генезе отдельных волн вызванного потенциала был описан механизм, обеспечивающий указанную двойную корреляцию.
В основе этого механизма лежало кольцевое движение нервных импульсов с «центральной станцией» в проекционной коре. Сначала возбуждение поступало от проекционной к ассоциативной коре. Для зрительных стимулов это были отделы височной коры, играющей важную роль в опознании стимулов, а для кожных раздражений – вторичные и третичные зоны соматосенсорной коры. Затем нервные импульсы приходили к структурам лимбико-гиппокампального комплекса и подкорковым центрам эмоций и мотиваций. До этой стадии поступательное движение возбуждения от рецептивных зон к исполнительным центрам полностью укладывалось в схему рефлекса. Однако процесс на этом не заканчивался: он включал еще возврат возбуждения в кору, включая и ее проекционные отделы (по системе диффузных проекций). Благодаря возврату возбуждения нервные импульсы, приходящие из мотивационных центров, накладывались в нейронах проекционной коры на следы сенсорного возбуждения. На этом этапе или несколько раньше в процесс включалась и лобная кора, что проявлялось в синхронизации рисунка ВП в интервале от 100 до 200 мс после стимула [Иваницкий, Стрелец, 1979]. Было высказано предположение, что указанные промежуточные компоненты ВП отражали синтез на корковых нейронах двух видов информации: наличной информации о физических характеристиках стимула и извлекаемых из памяти сведений о его значимости.
Самое интересное, однако, заключалось в том, что пиковая латентность этих волн ВП точно совпала со временем появления ощущения, измеренным ранее в психологических экспериментах [Froelich, 1929; Pieron, 1961; Бойко, 1964]. Следовательно, можно было предположить, что синтез двух видов информации – наличной и извлекаемой из памяти, – и составляет тот ключевой механизм, который лежит в основе ощущения как феномена уже не физиологического, а психического уровня (рис. 11.1).
Другими словами, был сделан шаг к преодолению барьера между двумя уровнями организации мозговых процессов, один из которых не сопровождается, а другой сопровождается субъективными переживаниями. Согласно предложенной концепции, последовательное поступление информации от рецепторов приводит к повторному движению возбуждения по указанному кольцу, обеспечивая постоянное сопоставление сигналов, приходящих из внешней и внутренней среды, что и составляет психический мониторинг происходящих изменений. Этот процесс осуществляется с периодом квантования приблизительно в 100–180 мс (обратим внимание, что это время сопоставимо с указанным в теории прожектора), составляющим минимальную длительность ощущений [Гольдбурт, Макаров, 1971; Blumenthal, 1977]. Позднее сходное кольцо возбуждения было описано М. Мишкиным [Mishkin, 1993] у обезьян при анализе стимульной информации и образовании памятного следа.
Идея о повторном входе возбуждения в нервные структуры как базисном механизме возникновения субъективного опыта была высказана в последние годы (независимо от нас) рядом авторов. Наиболее полно она была развита в трудах нобелевского лауреата (за исследование антител) Дж. Эделмена [Эделмен, 1981; Edelman, 1989], теория сознания которого получила достаточно широкую известность. По мнению этого автора, в основе возникновения субъективных феноменов лежит механизм повторного входа возбуждения в те же нейронные группы после дополнительной обработки информации в других группах или поступления сигналов из внешней среды, причем обратные связи могут соединять как анатомически близкие, так и удаленные структуры. Этот повторный вход (reentering) дает возможность сравнивать имевшиеся ранее сведения с изменениями, происшедшими в течение одного цикла.
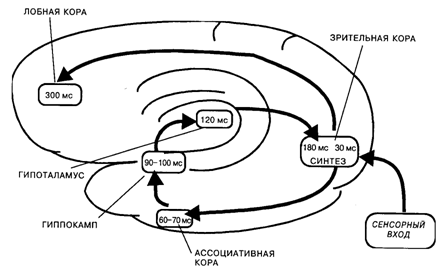
Рис. 11.1. Схема кольцевого движения возбуждения при возникновении зрительных ощущений. Ведущее звено в этом процессе – синтез информации о физических и сигнальных свойствах стимула на нейронах проекционной коры
Кроме того, сходная концепция сознания разрабатывается в нашей стране В. Сергиным [1994], который считает, что данный механизм лежит в основе феномена «внутреннего видения», составляющего суть сознания. Сходные гипотезы о возврате возбуждения в первичную кору как механизме возникновения зрительных и соматосенсорных ощущений были выдвинуты и другими авторами [Cauller, Kulics, 1991; Stoerig, Brandt, 1993; Desmedt, Tomberg, 1995].
Достаточно хорошо проработанная с концептуальной и нейрофизиологической точки зрения концепция сознания была выдвинута Дж. Греем [Gray, 1995]. В соответствии с этой концепцией, содержание сознания определяется активностью субикулярного (отдел гиппокампа) компаратора вместе с обратной связью от данного компаратора к тем наборам нейронов в перцептивной системе, которые снабжают вход данного компаратора с учетом результатов текущего процесса сравнения. Идея о важной роли лимбических структур в генезе психики согласуется и с данными о том, что эти структуры тесно связаны с эксплицитной памятью, узнаванием и вспоминанием [Mishkin et al., 1991]. В то же время вряд ли гиппокампу может принадлежать главная роль в высших функциях мозга. Именно поэтому представления о синтезе информации в коре как ведущем звене в механизмах психики, с нашей точки зрения, более предпочтительны, тем более, что они хорошо согласуются и с данными других авторов, приведенными ранее.
Для оценки предложенных концепций важное значение может иметь соотнесение временной шкалы мозговых процессов со временем субъективно переживаемых событий. По Дж. Эделмену [Edelman, 1989], время одного цикла прохождения возбуждения составляет 100– 150 мс. Если прибавить к этому время, необходимое для прихода сенсорных импульсов в кору, то в сумме получается время, достаточно близкое к тому, которое было найдено в наших экспериментах. Дж. Грей [Gray, 1995] считает, что в квантовании процессов сознания ведущую роль играют процессы, связанные с тета-ритмом, что дает следующее время: 1000/6=166 мс. О значении частот тета-ритма в данном аспекте в свое время писал и П.В. Симонов [1979]. В цитированной ранее работе Дж. Десмедта и К. Томберг [Desmedt, Tomberg, 1995] процесс синхронизации на частоте 40 Гц развивался в течение 100 мс после появления потенциала в первичной коре и до начала волны Р300. Попутно отметим, что синхронизации потенциалов на частоте 40 Гц рядом автором придается большое значение в механизмах сознания и образования зрительных образов [Kulli, Koch, 1991; Engel et al., 1991].
M. Познер и М. Ротбарт [Posner, Rothbart, 1994] показали, что при опознании стимулов вслед за первичной активацией задних отделов коры происходит вовлечение в функцию фронтальной коры с последующим возвратом возбуждения и реактивацией задней коры через 150 мс после ее первоначального возбуждения.
В исследованиях Б. Либета и др. [Libet et al., 1967] с регистрацией вызванных потенциалов на электрокожные стимулы с поверхности коры во время нейрохирургических операций было показано, что на слабые, подпороговые стимулы в коре регистрируются только ранние волны ответа с латентностью до 100 мс. При усилении раздражений в ВП появляются и более поздние колебания с латентностью 150 мс, что сопровождается появлением субъективных ощущений и практически совпадает с латентностью волн ВП, обнаруживших в наших исследованиях двойную корреляцию с перцептивными индексами.
Б. Баарс [Baars, 1993] на основе анализа психологической литературы приходит к выводу, что образы возникают в течение первых 200 мс после действия стимула, а в последующем (200–500 мс) происходит их категоризация.
Из сказанного можно сделать вывод о том, что идея повторного входа возбуждения и информационного синтеза в течение последних 20 лет была высказана независимо друг от друга рядом авторов, относящихся к разным школам и строившим свои заключения на основе разных данных. Очевидно, подобное совпадение не может рассматриваться как случайное. Скорее, оно свидетельствует о приближении к истинному пониманию тех принципов организации мозговых процессов, которые лежат в основе субъективных переживаний.
Приведенные ранее данные касались происхождения одного из наиболее простых психических явлений – ощущений. В связи с этим может возникнуть вопрос о том, насколько данные закономерности универсальны и может ли тот же принцип информационного синтеза быть распространен и на более сложные психические проявления, например на процесс мышления?
Механизмы мышления
Ответу на этот вопрос были посвящены наши исследования последних десяти лет. В работе ставилась задача изучить структуру корковых связей при решении различных мыслительных операций. При этом мы исходили из фундаментальных идей российской нейрофизиологической школы Н.Е. Введенского–А.А. Ухтомского о том, что связь между нервными структурами возникает на основе уравнивания ритмов их деятельности. М.Н. Ливановым [1972] и B. C. Русиновым [1969] было показано, что эти ритмы находят отражение в частотных параметрах электроэнцефалограммы (ЭЭГ), из чего последовал решающий вывод о том, что синхронизация частотных характеристик биопотенциалов мозга может явиться условием и индикатором внутрикорковой связи.
В развитие этих идей был создан метод, получивший название картирования внутрикоркового взаимодействия [Иваницкий, 1990, 1997]. В основе метода лежат представления о том, что наличие в спектрах ЭЭГ разных областей коры точно совпадающих частотных пиков является указанием на наличие в этих областях нейронных групп, работающих в одном ритме и, следовательно, функционально связанных друг с другом.
Испытуемым на экране монитора предъявлялись задачи на образное, пространственное и абстрактно-вербальное мышление. В первом случае субъект должен был опознать эмоцию на фотографии лица (использовались четыре базовые эмоции: радости, страха, гнева и горя, а также смешанные состояния), во втором – сравнить две геометрические фигуры с целью определения их идентичности или зеркальной симметрии. В качестве вербальных задач использовано решение анаграмм или выбор из четырех слов одного, относящегося к другой смысловой группе.
Было установлено, что простой и достаточно симметричный рисунок связей, характерный для состояния покоя, при умственной работе изменялся. Связи начинали сходиться к определенным областям коры, образуя как бы узлы или центры связей, названные фокусами взаимодействия. При этом топография фокусов взаимодействия оказалась специфичной для мыслительных операций разного знака. Так, при образном мышлении фокусы локализовались преимущественно в теменно-височных областях, а при абстрактно-вербальном мышлении – в лобных отделах коры. Пространственные задачи, включавшие элементы обоих видов мышления, характеризовались образованием фокусов на более ранних этапах в задних, а затем и в передних отделах коры [Сидорова, Костюнина, 1991; Иваницкий, Ильюченок, 1992; Николаев и др., 1996] (рис. 11.2).
Специализация полушарных функций как бы накладывается на описанные ранее закономерности. В исследованиях, в которых испытуемому давалось задание мысленно построить зрительный образ из ограниченного набора простых элементов, было показано, что у лиц с преобладанием первой сигнальной системы, по И. П. Павлову, фокусы взаимодействия локализовались преимущественно в правом, а у лиц с преобладанием второй, речевой системы – в левом полушарии. При этом на этапе нахождения образа фокусы были расположены в затылочных и височных (зона опознания) отделах полушарий, а на этапе конструирования образа – в лобной коре [Иваницкий и др., 1990]. Следует отметить, что нахождение решения при всех типах задач, даже если речевой ответ не требовался, сопровождалось (или определялось?) включением в функцию вербальной левой височной зоны.
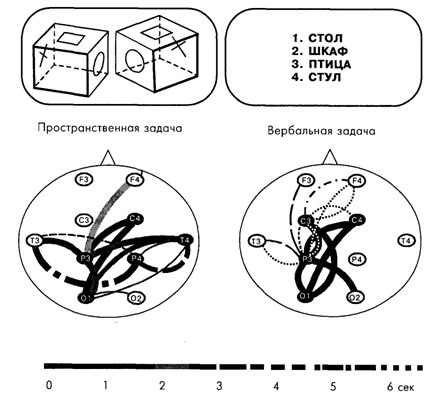
Рис. 11.2. Корковые связи в диапазоне частот бета-ритма при двух мыслительных операциях. Вверху представлены примеры задач; слева – связи при задаче на пространственное мышление, которая заключалась в сравнении двух геометрических фигур, предъявляемых на экране монитора, с целью определения их идентичности или зеркальной симметрии, что решалось путем мысленной ротации; справа – связи при задаче на вербальное мышление, которая состояла в том, что из четырех слов на экране монитора субъект должен был выбрать одно, относящееся к другой смысловой группе. На рисунке представлены статистически значимые связи при сравнении со зрительно-моторным контролем, включавшим предъявление изображений и двигательный ответ, но не требовавшим мыслительных операций. Толстые линии – связи на частоте 16–19 Гц., тонкие линии – связи на частоте 13–16 Гц. Шкала – время появления связи после предъявления задачи. Средние данные по группе из 43 человек
Обобщая эти данные, можно прийти к выводу о том, что важным признаком организации корковых связей при мышлении является их конвергенция к определенным центрам – фокусам взаимодействия. При этом связи, подходящие к фокусу, устанавливаются на разных частотах, собственно именно это обстоятельство и лежит в основе образования фокуса, так как связи на одной частоте образовывали бы однородную сеть, не имеющую центров. Можно предполагать, что каждая из связей приносит к центру из определенной области коры или подкорковых образований свою информацию. В фокусе эта информация может быть сопоставлена и перекомбинирована определенным образом. Основную функцию фокуса взаимодействия составляет, таким образом, информационный синтез, т.е. процесс, сходный с тем, который мы наблюдали в проекционной коре при возникновении ощущений.
Основное различие состоит в том, что место сенсорного сигнала здесь могла занять информация, хранящаяся в оперативной памяти (например, об условиях решаемой задачи), а ведущая роль в процессах информационного синтеза принадлежала не проекционной, как при возникновении ощущений, а ассоциативной коре. В фокусе оперативная информация сопоставлялась с информацией, извлекаемой из долговременной памяти, и сигналами, приходящими из мотивационных центров. Предполагается, что на основе происходящего в фокусе сопоставления и достигается конечная цель мыслительного процесса в виде нахождения решения. Субъективно все это переживается как процесс думания и нахождения ответа.
Гипотетическая структура фокуса взаимодействия состоит из групп нейронов с различными частотными характеристиками, настроенных на одинаковые с ними по частоте группы на периферии. Эти связи по своей природе должны быть двухсторонними, т.е. как прямыми, так и обратными: если две группы имеют одну частоту, то любая из них в равной степени способна и воспринимать, и передавать информацию связанной с ней группе (в зависимости от соотношения фаз колебаний). Одно кольцо при ощущении заменяется здесь, таким образом, как бы системой колец, замыкающихся на один центр.
Внутри фокуса группы нейронов должны быть объединены связями, образованными на ином принципе: так как они работают на разных частотах, принцип изолабильности здесь не применим. Очевидно, это должны быть жесткие связи, основанные на структурных изменениях в синапсах. Данные связи эффективны в любой фазе цикла возбудимости нейрона или нейронного осциллятора, за исключением фазы абсолютной рефрактерности (рис. 11.3). Идея об обеспечении психической функции за счет сочетания жестких и гибких звеньев была впервые высказана Н.П. Бехтеревой [1980].
Изложенные представления о фокусах взаимодействия и их функциональном значении хорошо согласуются с данными А. Дамасио [Damasio, 1994], который считает, что в обеспечении высших психических функций ведущую роль играют так называемые зоны конвергенции, которые принимают и синтезируют информацию, поступающую из других отделов коры и подкорковых образований.
Все сказанное дает возможность объединить данные, полученные при изучении механизмов восприятия и мышления, единым принципом информационного синтеза как мозговой основы возникновения нового качества в виде субъективных переживаний.
Дата добавления: 2019-04-03; просмотров: 593;