Зрительные обобщения и представления
Подлинная рецепция, истинное восприятие предметных компонентов среды как таковых возможны лишь на основе достаточно развитой способности к анализу и обобщению, ибо только это позволяет полноценно узнавать постоянно меняющие свой вид (и другие свойства) предметные компоненты среды.
Исследования, проведенные на разных видах позвоночных (помимо круглоротых), показали, что все они способны к предметному восприятию, в частности к восприятию форм. Уже на этом основании можно заключить, что все позвоночные находятся на стадии перцептивной психики. Однако внутри этой стадии наблюдаются существенные различия между низшими и высшими позвоночными.
Так, рыбы способны в эксперименте отличать геометрические фигуры, например квадрат от треугольника, независимо от меняющейся величины этих фигур. Но достаточно в контрольном опыте перевернуть треугольник (острием вниз) или квадрат (придать ему положение ромба), как рыба перестает узнавать эти фигуры. Следовательно, восприятие и узнавание формы оказываются здесь ограниченными, недостаточно гибкими. Отсутствует способность к быстрому переносу сформировавшегося зрительного представления на основе обобщения. Млекопитающие же вполне способны к такому обобщению и легко узнают треугольник или другую геометрическую фигуру любой величины в любом положении. Существенно превосходят рыб также птицы. Правда, и курица, по некоторым данным, не узнает перевернутый треугольник, которыйдо этого безошибочно отличала от другой геометрической фигуры. Вместе с тем курица способна к значительным обобщениям, опознавая, например, геометрические фигуры независимо от их цвета. Более того, будучи обучена выбирать сплошной треугольник, она также реагирует на него, когда он предъявляется ей в виде штриховки или даже лишь намечен тремя точками! Так же как и рыбы, птицы способны распознавать относительную величину фигур и так же ошибаются на основе зрительных иллюзий.
Способность к зрительному обобщению у многих позвоночных экспериментально доказана. Особенно в отношении внешних их представителей было показано, что они в состоянии даже в очень сложных ситуациях вычленять существенные детали в воспринимаемых объектах и узнавать эти объекты в сильно измененном виде. Отсюда напрашивается вывод о существовании у позвоночных достаточно сложных общих представлений. Представления чрезвычайно важны для выживания. В них закрепляется в общем виде индивидуальный опыт, что позволяет животному легче ориентироваться в существенно меняющейся окружающей среде при временном отсутствии жизненно важных раздражителей. Подобные зрительные обобщения обнаруживаются, в частности, в явлениях «экстраполяции», описанных Л.В.Крушинским, которые проявляются в том, что в ряде случаев животные способны, проследив за движением какого-либо предмета, ориентироваться затем на ожидаемый путь его перемещения. Следовательно, учитывается предстоящее передвижение объекта после его исчезновения. В проведенных им опытах птицы и млекопитающие помещались перед туннелем, имевшим посредине разрыв, через который можно было наблюдать за движением перемещающейся в туннеле кормушки с приманкой. В других опытах применялась ширма со щелью. Некоторые из подопытных животных (врановые, хищные) обегали затем туннель или ширму по направлению движения приманки и ожидали ее в месте появления. На такое «предвосхищающее» поведение оказались, однако,способными лишь животные, имеющие в естественных условиях дело с прячущейся добычей, которую приходится выслеживать или преследовать, зачастую обгоняя ее, или же сами прячущие корм про запас. Как уже раньше указывалось, мы имеем здесь дело с широко распространенной среди высших позвоночных способностью к нахождению обходного пути. При этом важно отметить, что в любом случае, когда животное огибает непрозрачную преграду или даже просто отворачивается от целевого объекта, оно на время теряет последний из виду. Более того, каждое представление по самой природе своей требует отсутствия ранее воспринятых объектов.
Убедительные доказательства тому, что обобщенные зрительные образы в форме представлений широко распространены среди позвоночных, дали эксперименты, проведенные методом отсроченных реакций (исследования В.С.Хантера, Р.Йеркса, О.Л.Тинклпоу, Ф.Бойтендайка, В.Фишеля и др.). В Советском Союзе такие эксперименты проводились Н.Ю.Войтонисом, И.С.Беритовым, Г.З.Рогинским и др. Обычная схема проведения подобных опытов такова: на виду у животного прячется корм, который, однако, позволяется ему отыскать лишь некоторое время спустя. Обычно подопытное животное должно произвести выбор между несколькими местами, где может находиться спрятанная приманка. При успешном решении задачи имеет место реакция на отсутствующий стимул, что возможно лишь при наличии чувственных представлений.
Опыты по методу отсроченных реакций с успехом ставились над различными позвоночными, особенно же часто над обезьянами. Весьма эффектными были опыты, которые проводил Тинклпоу, когда на глазах у шимпанзе . прятались фрукты (бананы), которые затем незаметно для животного подменялись значительно менее привлекательным кормом — салатом или капустой. Найдя спустя положенное время зелень, обезьяна медлила брать ее и продолжала, иногда с визгом, поиски. Последние продолжались до 33 секунд, в то время как в контрольном опыте, когда обезьяне с самого начала показывался салат, который затем не подменялся, поиски продолжались лишь 3 секунды и обезьяна спокойно съедала его. Напрашивается вывод, что обезьяна ищет виденный ею вначале объект, руководствуясь зрительным представлением, которое, по Тинклпоу, отражает также определенные количественные и качественные показатели.
Войтонис на основании своих экспериментов также пришел к выводу, что обезьяны (макаки и шимпанзе) реагируют не на кормушку, содержащую приманку, а на корм как таковой, точнее, на специфический вид корма. То же самое относится и к собакам, но не к рыбам (опыты Бойтендайка и Фишеля).
Наличие у позвоночных (а возможно, и у некоторых высших беспозвоночных) представлений, выражающихся в отсроченных реакциях и способности к нахождению обходных путей (включая и явления экстраполяции), придает их поведению исключительную гибкость и намного повышает эффективность их действий на поисковых этапах поведенческих актов. Вместе с тем не следует переоценивать эти способности, ибо они не обязательно свидетельствуют о высоком уровне психического развития. Основываясь на развитой памяти, они встречаются у позвоночных на разных филогенетических ступенях в зависимости от образа жизни животного: наряду с животными, прячущими в разных местах запасы, эти способности свойственны хищникам, охотящимся на труднодосягаемых, прячущихся или лишь эпизодически появляющихся на виду животных.
На эту биологическую обусловленность реакции в отсутствии стимула, который до этого ее обусловливал, указал еще полвека тому назад советский зоолог и зоопсихолог Д.Н.Кашкаров на примере кошки, которая «стережет мышь и видит, что мышь появляется из норы. Мышь исчезает прежде, чем кошка начнет реагировать. Однако кошка встает и идет к норе. Здесь не может быть вопроса о том, что в момент реакции определяющий стимул отсутствует... Правильный выбор между тремя норами кошка могла бы сделать лишь на основании непосредственно предыдущегопоявления мыши, т.е. на основе представления о последней»3. Что же касается обезьян, то, как видно из упомянутых исследований, мы явно имеем здесь дело с более сложными комплексными процессами.
Способность высших позвоночных к предметному восприятию, способности к анализу и обобщению, а тем самым к формированию представлений являются важной предпосылкой образования сложных навыков, которые уже рассматривались раньше (см. ч. I) и которые составляют основное содержание накопления индивидуального опыта не только в сенсорной, но и в эффекторной сфере, точнее сказать, в моторно-сенсорной сфере, действующей как единая система.
Общение
У высших позвоночных особой сложности достигают и процессы общения. Как и у других животных, средства коммуникации включают у них элементы различной модальности — ольфакторные, тактильные.
Ольфакторная сигнализация, т.е. передача информации другим особям химическим путем, преобладает в территориальном поведении, особенно при маркировке местности у хищных и копытных. Для этого служат специальные железы, расположенные в различных участках кожи и выделяющие специфический пахучий секрет. Последний распространяется по воздуху или наносится на различные предметы (стволы и ветки растений, камни и т.д.). Пахучие метки производятся также с помощью испражнений. Выделениями пахучих желез маркируется и след, что способствует нахождению друг друга особями одного вида. Однако во всех этих случаях общение существенно не отличается от того, что мы наблюдаем, скажем, у насекомых.
Видотипичные, инстинктивные компоненты поведения позвоночных, служащие для акустического и оптического общения между животными, как правило, ритуализованы. Оптическое общение осуществляется прежде всего с помощью выразительных поз и телодвижений, о чем уже говорилось выше. Видоспецифические отличия выражаются часто в малозаметных человеку, но четко генетически фиксированных деталях, вызывающих совершенно специфические, зачастую ритуализованные реакции у особей того же вида. Особенно часто такие различия наблюдаются у близкородственных животных, причем прежде всего в сигнальных системах, связанных с размножением. Здесь они служат одним из факторов биологического разграничения видов.
Конкретные формы оптического общения отличаются у высших позвоночных большим разнообразием и дифференцированностью. Нередко они выражаются в специфических «диалогах» между двумя или несколькими особями, т.е. во взаимной демонстрации частей тела или поз. Особенно это относится к «ритуалам» запугивания или «импонирования». «Импонирование» выражает превосходство одной особи по отношению к другой, имеет и оттенок угрозы и вызова, но, в сущности, не носит агрессивного характера (в отличие от подлинного «запугивания», за которым часто следует нападение). Часто самцы «импонируют» самкам во время «ухаживания» за ними. Подчиненная особь отвечает на «импонирование» позой «покорности», чем окончательно исключается возможность столкновений между животными.
У млекопитающих оптическое общение часто сочетается с ольфакторным (рис. 43), так что выделение систем общения по отдельным модальностям является у этих животных в большой степени условным.
Это относится и к акустическому общению, в качестве примера которого следует, конечно, прежде всего напомнить о криках и песнях птиц. Хотя человеческое ухо неспособно улавливать все издаваемые птицами звуки (значительная часть их находится в ультразвуковом спектре ниже 50 кГц), мы поражаемся огромному разнообразию их голосовых реакций. К последним необходимо добавить и неголосовые звуки, как, например, свист крыльев при полете, постукивание дятла по стволу деревьев и т.д. Видоспецифичность таких звуков делает их пригодными для общения.
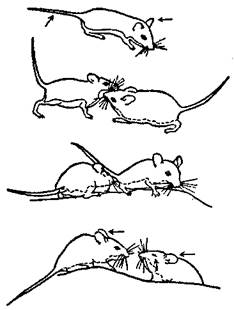
Рис. 43. Оптико-ольфакторное общение у грызунов (род Peromyscus). Стрелками обозначены места взаимного обнюхивания (по Эйзенбергу)
Для биологически адекватного реагирования на все эти звуковые сигналы существенной является видоспецифическая настройка слуховой системы на определенную часть спектра. Как показал советский исследователь биоакустики птиц В.Д.Ильичев, важнейшую роль играют в этой настройке периферические отделы слухового анализатора, выполняющие роль видотипических биологических фильтров. С их помощью осуществляется и специальная «подстройка» слуховой системы на особенно важные для особи звуки. Таким образом, здесь четко выступает характерная для всего инстинктивного поведения взаимообусловленность и единство врожденных функциональных (физиологических и этологических) и морфологических элементов.
Зоосемантика звуковых сигналов птиц охватывает практически все сферы их жизнедеятельности. Эти сигналы служат опознаванию особи (в частности, ее видовой принадлежности), оповещению других особей о физиологическом состоянии экспедиента (сюда относится, например, «попрошайничество») или об изменениях в окружающей среде (появление врага или, наоборот, кормового объекта и т.п.). При этом богатые зоопрагматические средства птицы позволяют экспедиенту передавать перцепиенту весьма детализованную информацию. Так, грач, подлетающий с кормом к гнезду, оповещает об этом насиживающую самку криком, могущим иметь четыре разных оттенка, на которые самка также отвечает разными звуками. Эти оттенки голосовых реакций птиц подчас настолько тонки, что не воспринимаются человеческим ухом.
Очень большое значение имеют не только четкие межвидовые различия акустического общения, но и индивидуальные, по которым особи узнают друг друга (самец — самку, птенцы — родителей и друг друга, члены стаи — друг друга и т.п.). Интересно, например, что, по данным В.Г.Торпа, в ряде случаев совместно поющие партнеры откликаются только на голос друг друга, но не реагируют на пение других особей. Особенно индивидуально различными являются территориальные крики птиц, оповещающих о занятости участка.
Большого разнообразия звуки достигают и у таких птиц, как куриные. Это обусловлено жизнью диких кур в труднопросматриваемых густых кустарниковых зарослях, где условия для оптического общения особенно неблагоприятны. Наиболее громкие и «впечатляющие» звуки куриных птиц — это крики петухов. Их слышимость (для человеческого уха) достигает 2 километров. В отличие, например, от звуков, с помощью которых петух подзывает кур к найденному им корму, кукареканье производится однократно, но зато дольше и на более высоких частотах. Это относится и к звуковым сигналам тревоги (например, при обнаружении врага), испуга или угрозы. Чем выше тональность, громкость и длительность этих сигналов, тем сильнее их эффект. Свое «кукареку» петух кричит на самых «высоких нотах», но всегда одинаковым образом: отклонения не превышают полтона.
Кукареканье — это своего рода вызов другим петухам, и те отвечают ответными криками, правда, лишь в том случае, если соперники — петухи одного «ранга». Если же в пределах видимости закричит петух, занимающий подчиненное положение, то господствующий попросту атакует его, «не удостаивая» ответным криком. Когда же перекликаются петухи, разделенные большим расстоянием и не видящие друг друга, то это типичные территориальные крики, т.е. акустическая маркировка местности, оповещение о занятости участков стайками кур — петухом и его «гаремом». Большое же биологическое значение крика заключается в том, что он предотвращает или хотя бы уменьшает частоту петушиных боев, а вместе с тем обеспечивает захват, освоение и защиту участков, а в конечном итоге расселение вида, но безвредным путем, ибо дело ограничивается одним запугиванием. Петух-соперник заранее предупреждается о том, что данный участок уже занят и ему придется искать другое пригодное для поселения место. И только в том случае, если все такие места уже заняты, начнутся бои за овладение участками. В этом случае победитель — животное более сильное не только в физическом, но и в психическом отношении — станет продолжателем рода, и это, конечно, тоже выгодно для процветания вида, для его прогрессивной эволюции. Аналогичную роль играет у других птиц пение. У домашних же кур крестьянский двор и прилегающие места остаются для петуха его участком, о занятии которого он непрестанно напоминает криком, хотя это поведение уже потеряло свое биологическое значение, поскольку человек обеспечивает все потребности домашних животных и управляет эволюцией вида. Надо думать, что чрезвычайная близость расположения участков домашних петухов заставляет их интенсивнее кукарекать, чем это имело бы место в дикой природе, так как в селе популяция кур «сверхплотная».
Мы не останавливаемся на звуковом общении млекопитающих, так как у них господствуют те же закономерности, что и у птиц.
Дата добавления: 2017-02-20; просмотров: 470;