Электрические проявления активности головного мозга.
Еще в 1875 г. Ричард Кейтон (Caton) показал, что с помощью электрода, приложенного к поверхности мозга животного, можно зарегистрировать электрическую активность в виде волн. Правдич-Неминский в 1925 г. продемонстрировал возможность отведения потенциалов через интактный череп.
Первые отведения от головного мозга человека произвел Ганс Бергер (Berger) в 1924 г.; опубликовано это было в 1929 г. До 1938 г. Бергер опубликовал около 20 работ под одним названием: «Об электроэнцефалограмме человека».
Суммарная биоэлектрическая активность мозга.
Если показатели биоэлектрической активности мозга регистрируются через микроэлектроды, то они отражают активность локального (до 100 мкм в диаметре) участка мозга и называются фокальной активностью.
Когда электрод располагается в подкорковой структуре, то регистрируемая через него активность называется субкортикограммой. Если электрод располагается в коре (или на поверхности коры) мозга – электрокортикограммой (ЭКоГ).
При расположении электрода на поверхности кожи головы, регистрируется суммарная активность как коры, так и подкорковых структур – электроэнцефалограмма (ЭЭГ).
Все виды активности мозга сопровождаются определенными ритмами электрических колебаний, которые подвержены усилению и ослаблению (рис. 17; рис. 18).
У человека в покое при отсутствии внешних раздражений чаще всего на ЭЭГ преобладает альфа-ритм: частота 8-13 Гц, амплитуда 50 мкВ.
Переход к активной деятельности приводит к смене альфа-ритма на более быстрый бета-ритм, имеющий частоту колебаний 14-30 Гц, амплитудой в 25 мкВ. Бета-подобная активность наблюдается также во время «парадоксальных» фаз «быстрого» сна (сопровождающихся быстрыми движениями глаз).
Переход от состояния покоя к состоянию сосредоточенного внимания, или, наоборот, ко сну (дремотному состоянию) сопровождается развитием тэта-ритма: 4-8 Гц, амплитуда 100 мкВ.
Дельта-ритм – частота 0,5-3,5 Гц, амплитуда 200-300 мкВ, регистрируется во время глубокого «медленного» сна.
Механизмы возникновения ЭЭГ и ЭКоГ, снимаемой с поверхности коры, одни и те же. Однако амплитуда зубцов ЭЭГ ниже вследствие электрического сопротивления тканей, находящихся между поверхностью коры мозга и регистрирующими электродами. Частота волн ЭЭГ также несколько меньше, так как в связи с большей удаленностью электродов от потенциальных генераторов электрической активности регистрируется деятельность более обширных участков коры и быстрые колебания потенциалов взаимокомпенсируются.
Суммарные биоэлектрические процессы в коре мозга тесно связаны с динамикой зарядов мембран нейронов, глии, с процессами в синапсах, дендритах, аксонном холмике, в аксоне.
Первоначальные гипотезы связывали процесс формирования ЭКоГ (а, соответственно и ЭЭГ) преимущественно с суммацией ПД. Однако длительность ПД нейронов коры составляет 0,5 – 2 мс.
В настоящее время считается, что ЭКоГ отражает главным образом постсинаптическую активность нейронов коры. ПСП нейронов коры длительнее, чем у мотонейронов. Восходящая фаза ВПСП длится несколько мс, а нисходящая – 10-30 мс. ТПСП корковых нейронов еще длительнее – 70-150 мс. Несколько упрощая ситуацию, можно сказать, что положительное отклонение потенциала на поверхности коры вызывается тормозными постсинаптическими потенциалами в поверхностных слоях коры, либо возбуждающими постсинаптическими потенциалами в ее глубинных слоях, а отрицательное отклонение – противоположными причинами.
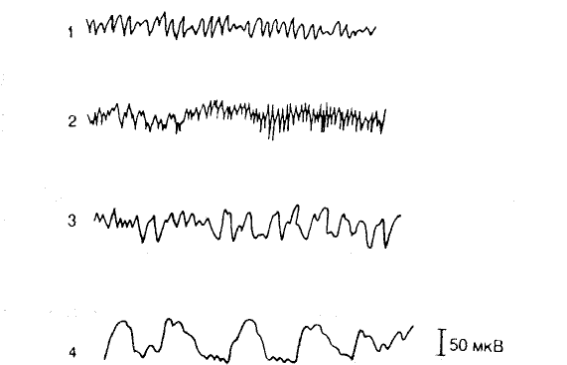
Рис. 17. Основные ритмы ЭЭГ.
1 – альфа-ритм; 2 – бета-ритм; 3 – тета-ритм; 4 – дельта-ритм.
Ритмичность активности коры индуцируется главным образом активностью подкорковых структур. В частности, для проявления альфа-активности особенно важен таламус. Таламические ритмоводители (пейсмекеры) за счет своих возбуждающих и тормозящих связей способны генерировать и поддерживать ритмическую активность. Эта активность модифицируется связями таламуса с другими структурами мозга. Особенно выраженным синхронизирующим (генерирующим ритм) и десинхронизирующим (подавляющим ритм) действием на таламус обладает ретикулярная формация.
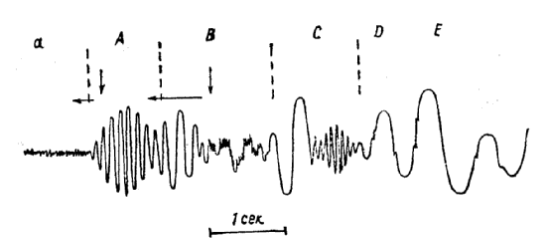
Рис. 18. Стадии ЭЭГ.
а – возбужденное состояние, проявляется в виде бета- и более быстрых ритмов малой амплитуды; А – состояние спокойного бодрствования при закрытых глазах в темноте, чаще всего характеризуется хорошо выраженным альфа-ритмом; В – легкое полудремотное состояние, вместо альфа-ритма появляются нерегулярные колебания разной частоты, иногда стойкий тета-ритм, иногда смесь дельта- и тета-волн; С – начальная фаза сна, появляются дельта-волны большой амплитуды среди которых возникают вспышки веретенного ритма (13,5 – 14 Гц); D и E – стадии более глубокого «медленноволнового» сна с хорошо выраженными крупными дельта-волнами, по мере углубления сна происходит некоторое уменьшение частоты и амплитуды колебаний.
Вызванные потенциалы (ВП).
Когда на фоне покоя (или другого состояния) предъявляется быстрое нарастающее раздражение, на ЭЭГ регистрируются ВП – т.е. синхронная реакция множества нейронов данной зоны на стимул. Компоненты ВП, количество и характер колебаний зависят от адекватности стимула относительно зоны регистрации ВП. ВП может состоять из первичного или из первичного и вторичного ответов (рис. 19). 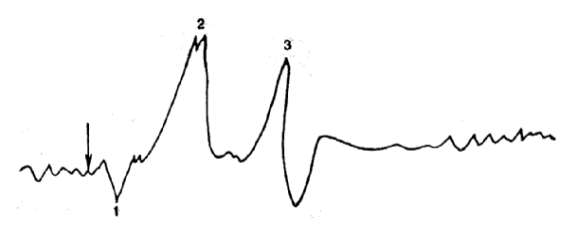 Рис. 19. Вызванный потенциал в зрительной области коры на световое раздражение.
Рис. 19. Вызванный потенциал в зрительной области коры на световое раздражение.
1 – позитивное колебание; 2 – первичное негативное колебание; 3 – вторичные колебания.
Первичные ответы регистрируются в первичных зонах коры анализатора при адекватном для данного анализатора стимуле. Характеризуются коротким латентным периодом, двухфазностью колебания: вначале положительное, затем отрицательное. Формируются за счет кратковременной синхронизации активности близлежащих нейронов.
Вторичные ответы более вариабельны по латентным периодам, длительности и амплитуде, охватывают более обширную корковую область. Они чаще возникают на сигналы, не только адекватные для данного анализатора, но и имеющие определенную смысловую нагрузку.
Значение регистрации ВП заключается в возможности судить по этим данным о сохранности периферических и подкорковых сенсорных путей.
Постоянные потенциалы коры головного мозга.
Обычно между поверхностью коры и нижележащим белым веществом или между корой и удаленным от нее индифферентным электродом существует постоянная поверхностно-отрицательная разность потенциалов порядка нескольких мВ. Эти постоянные потенциалы также изменчивы, но частота их колебаний значительно ниже, чем у ЭКоГ. Например, во сне потенциал поверхности коры становится положительным, а при пробуждении или увеличении поведенческой активности – более отрицательным. Местная или генерализованная судорожная импульсация, нарушения транспорта дыхательных газов также приводят к характерным изменениям постоянного потенциала.
Общепринятого мнения о происхождении постоянных потенциалов коры нет. Вероятно, что сдвиги в отрицательную сторону обусловлены деполяризацией апикальных дендритов в I и II слоях, вызванной активностью неспецифических таламических афферентов. Косвенно на постоянные потенциалы могут влиять глиальные клетки.
Дата добавления: 2017-08-01; просмотров: 1000;