Перенос генетического материала бактерий
Обмен генетическим материалом у бактерий осуществляется путем генетических рекомбинаций. Под генетической рекомбинацией подразумевают взаимодействие между двумя геномами, которое приводит к образованию рекомбинаций ДНК и формированию дочернего генома, сочетающего гены обоих родителей. Особенности рекомбинаций у бактерий определяются отсутствием истинного полового процесса и мейоза у прокариот и гаплоидным набором генов. В процессе рекомбинации бактерии условно делятся на клетки-доноры, которые передают генетический материал, и клетки-реципиенты, которые этот материал воспринимают. В клетку-реципиент проникает не вся, а только часть хромосомы клетки-донора, т.е. один или несколько генов. Образуется только один рекомбинант, генотип которого представлен в основном генотипом реципиента с включением фрагментов хромосомы донора.
Рекомбинация может быть гомологичной, при которой в процессе разрыва и воссоединения ДНК происходит обмен между участками ДНК, обладающими высокой степенью гомологии. Встречается также сайт-специфическая рекомбинация, которая происходит только в определенных участках (сайтах) генома и не требует высокой степени гомологии ДНК, например включение плазмиды в хромосому бактерии. Передача генетического материала между бактериями осуществляется 3-мя механизмами: конъюгацией, трансдукцией и трансформацией (рис. 31).
Коньюгация – это перенос генетического материала путем прямого контакта между двумя клетками. Необходимым условием конъюгации является наличие в клетке-доноре трансмиссивной плазмиды. Трансмиссивные плазмиды кодируют половые пили, образующие конъюгационную трубочку между клеткой-донором и клеткой-реципиентом, по которой плазмидная ДНК передается из клетки-донора в клетку-реципиент. В результате такого переноса клетка-реципиент получает донорские свойства. Интегративной трансмиссивной плазмидой является F-фактор. Клетки-доноры, обладающие F-фактором, обозначаются как F+-клетки, а клетки-реципиенты, не имеющие F-фактора – F– -клетки. Если F-фактор встраивается в хромосому клетки-донора и начинает функционировать в виде единого с хромосомой трансмиссивного репликона, то хромосома донора приобретает способность передаваться в клетку-реципиент. Донорские клетки, имеющие встроенный в хромосому F-фактор,
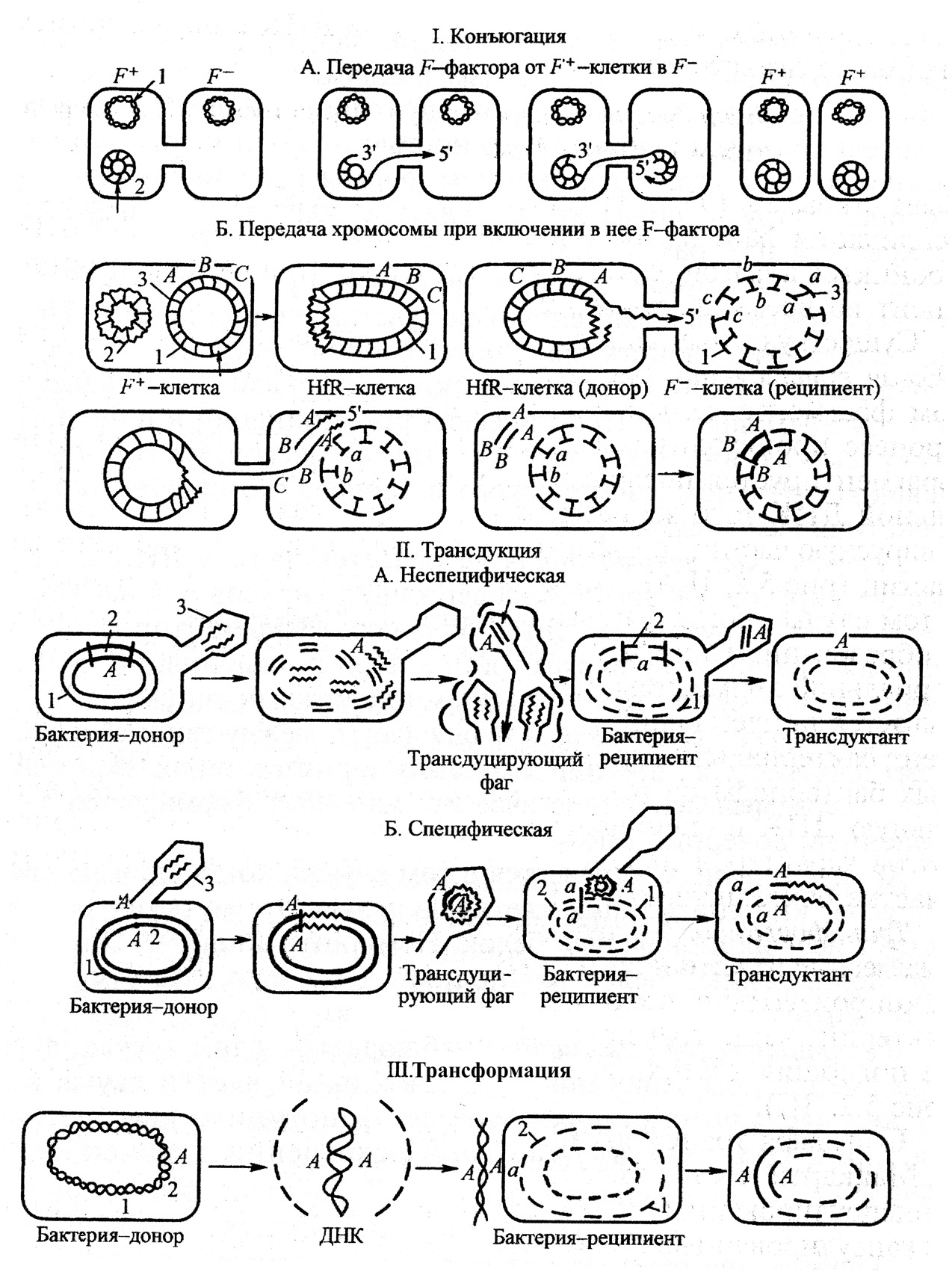
|
Рис. 31. Передача генетической информации у бактерий. I: 1 – бактериальная хромосома; 2 – F-фактор; 3 – гены, участвующие в рекомбинации; АВС – генотип донора; abc – генотип реципиента. II: 1 – бактериальная хромосома; 2 – гены, участвующие в рекомбинации: А – переносимый фагом ген; а – генотип реципиента; 3 – фаг. III: 1 – бактериальная хромосома; 2 – гены, участвующие в рекомбинации: А – генотип донора; а – генотип реципиента. (Микробиология и иммунология.-Под ред. Воробьева А.А. - М., 1999).
называются Hfr-клетками. Хромосомная ДНК реплицируется, одна цепь копии хромосомы переносится в реципиентную F– -клетку, тогда как другая остается в Hfr+-клетке, т.е. донор сохраняет свое генетическое постоянство.
Передача генетического материала при конъюгации начинается с расщепления ДНК в районе локализации F-фактора. Одна нить донорской ДНК передается через конъюгационный мостик в клетку-реципиент. Процесс сопровождается достраиванием комплементарной нити до образования двунитевой структуры. Переданная в реципиентную клетку и достроенная до двунитевой структуры, нить ДНК рекомбинирует с гомологичным участком реципиентной ДНК с образованием стабильной генетической структуры. Биологическая значимость конъюгации хорошо видна на примере распространения резистентности бактерий к антибиотикам. Устойчивость к антибиотикам бактерия может получить в результате мутации, что происходит 1 раз на каждые 106 клеточных делений. Однако, однажды изменившись, генетическая информация может быстро распространяться среди сходных бактерий посредством конъюгации, поскольку каждая третья из близкородственных бактерий способна именно к этому типу генетического переноса.
Трансформация – передача генетической информации через выделенную из клетки-донора ДНК. Процесс трансформации может произвольно происходить в природе у некоторых видов бактерий, чаще грамположительных, когда ДНК, выделенная из погибших клеток, захватывается реципиентными клетками. Как правило, любая чужеродная ДНК, попадающая в бактериальную клетку, расщепляется рестрикционными эндонуклеазами; но при некоторых условиях такая ДНК может быть интегрирована в геном бактерии. По происхождению ДНК может быть плазмидной либо хромосомной и нести гены, трансформирующие реципиента. Подобным путем процессы трансформации могут распространять гены, кодирующие факторы вирулентности, среди бактериальных популяций; однако в обмене генетической информацией трансформация играет незначительную роль.
Трансформация служит хорошим инструментом для картирования хромосом, поскольку трансформированные клетки включают различные фрагменты ДНК. Определение частоты одновременного приобретения двух заданных характеристик (чем ближе расположены гены, тем более вероятно, что они оба включатся в один и тот же участок ДНК) дает информацию о взаиморасположении соответствующих генов в хромосоме. Перенос экстрагированной ДНК является основным методом генной инженерии, используемым при конструировании рекомбинантных штаммов с заданным геномом.
Трансдукция – передача бактериальной ДНК посредством бактериофага. В процессе репликации фага внутри бактерий фрагмент бактериальной ДНК проникает в фаговую частицу и переносится вместе с ней в бактерию-реципиент. При этом фаговые частицы как правило дефектны, они теряют способность к репродукции. Так как трансдуцируются лишь небольшие фрагменты ДНК, вероятность рекомбинации, затрагивающей какой-то определенный признак, очень мала: она составляет от 10-6 до 10-8. Существуют три типа трансдукции: неспецифическая (общая), специфическая и абортивная.
Общая (неспецифическая) трансдукция – перенос бактериофагом фрагмента любой части бактериальной хромосомы. В клетке, инфицированной бактериофагом, в ходе сборки дочерней популяции в головки некоторых фагов может проникнуть фрагмент бактериальной ДНК или плазмиды либо вместе с вирусной ДНК, либо вместо нее. Этот процесс происходит вследствие того, что бактериальная ДНК фрагментируется после фаговой инфекции и кусочек бактериальной ДНК того же размера, что и фаговая ДНК, проникает в вирусную частицу с частотой приблизительно 1 на 1000 фаговых частиц. При такой форме трансдукции в клетки-реципиенты могут быть внесены практически любые гены. Феномен неспецифической трансдукции может быть использован для картирования бактериальной хромосомы.
Специфическая трансдукция наблюдается в том случае, когда фаговая ДНК интегрирует в бактерию с образованием профага. При исключении ДНК фага из бактериальной хромосомы в результате случайного процесса захватывается прилегающий к месту включения фаговой ДНК фрагмент бактериальной хромосомы. Так как большинство умеренных фагов интегрируют в бактериальную ДНК в специфических участках, для таких бактериофагов характерен перенос в клетку-реципиент определенного участка бактериальной ДНК донора. Специфическая трансдукция может служить механизмом переноса вирулентных генов среди бактерий при условии, что эти гены локализованы в непосредственной близости от мест интеграции профага.
Наиболее характерным примером служит трансдукция, осуществляемая фагом λ. Обычно он трансдуцирует определенные гены: gal (кодирует синтез галактозы) и bio (кодирует синтез биотина). При переходе в состояние профага фаг λ включается в определенный участок хромосомы бактерии-хозяина – между генами gal и bio. Отделение фаговой ДНК от бактериальной хромосомы может происходить неточно и какой-то фрагмент ее останется в хромосоме, а близко расположенные гены будут захвачены фаговой ДНК. В случае заражения трансдуцирующим фагом клеток, дефектных по определенному гену, например gal-, может произойти рекомбинация с заменой собственного дефектного гена бактерии интактным трансдуцированным геном с образованием рекомбинанта (трансдуктанта) gal+.
Абортивная трансдукция. При абортивной трансдукции внесенный фрагмент ДНК донора не встраивается в хромосому реципиента, а остается в цитоплазме и там самостоятельно функционирует. Впоследствии он передается одной из дочерних клеток (т.е. наследуется однолинейно) и затем теряется в потомстве.
Дата добавления: 2016-08-07; просмотров: 1736;