КЛИНИЧЕСКАЯ АНАТОМИЯ ВЕСТИБУЛЯРНОГО АНАЛИЗАТОРА
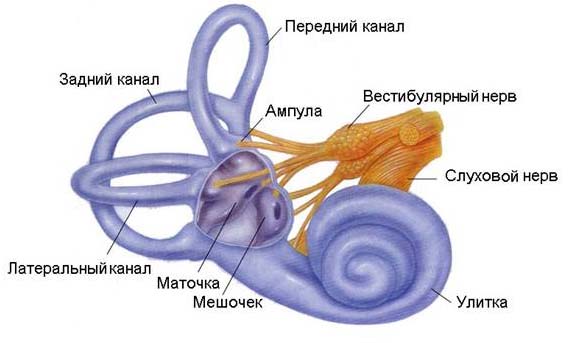 В пирамиде височной кости помещается орган, который за свою форму получил название «лабиринт». Структурные образования лабиринта называются преддверно-улитковым органом (рис. 16). В нем различают вестибулярный и улитковый отделы.
В пирамиде височной кости помещается орган, который за свою форму получил название «лабиринт». Структурные образования лабиринта называются преддверно-улитковым органом (рис. 16). В нем различают вестибулярный и улитковый отделы.
Улитковый отдел вместе с находящимися в нем чувствительными волосковыми клетками представляет собой периферический отдел слухового анализатора. Вестибулярный отдел является периферической частью вестибулярного анализатора, осуществляющего координацию позы (статики) и походки, движений (кинетики).
В периферических структурных образованиях вестибулярного анализатора различают две части: преддверие и систему полукружных каналов (рис. 17). В их костных образованиях расположен перепончатый лабиринт, содержащий чувствительные рецепторные клетки вестибулярного анализатора. В углублениях преддверия содержатся рецепторные структуры маточки (utriculus) и мешочка (sacculus), нейроэпителиальные клетки которых воспринимают прямолинейные (тангенциальные) ускорения, изменение положения головы и туловища в пространстве, силу притяжения и центробежную силу. В трех полукружных каналах расположены рецепторы, чувствительные к угловым (радиальным) ускорениям.
Схема анатомо-топографического расположения полукружных каналов, маточки и мешочка преддверия представлена на рис. 18.
Благодаря этим нейроэпителиальным образованиям, их рефлекторным связям со зрительными, тактильными, проприоцептивными и другими афферентными системами организма обеспечивается точная координация движений, четкое представление о положении тела в пространстве. Преддверие, содержащее отолитовый аппарат, филогенетически является более древним образованием, чем полукружные каналы.
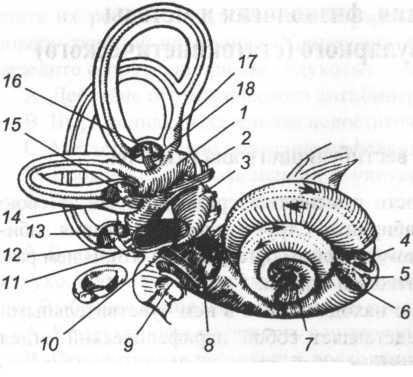
Рис. 16. Преддверно-улитковый орган (по B.I. Meloni, 1957): 1 — столбик; 2 — эндолимфатический проток; 3 — мешочек; 4 — барабанная лестница; 5,8 — улитковый проток; 6 — лестница преддверия; 7 — улитковый ход; 9 — окно улитки; 10 — мыс; 11 — окно преддверия; 12, 14, 18 — задний, боковой и передний полукружные каналы с соответствующими полукружными протоками; 13, 15, 16 — перепончатые ампулы полукружных каналов; 17 — общая перепончатая ножка

Рис. 17. Схема строения лабиринта (по Н. Frenzel, 1955): 1 — мешочек со статокониями, которые в нем содержатся; 2 — маточка; 3 — полукружные каналы; 4 — эндолимфатический проток; 5 — соединительный проток; 6 — улитка
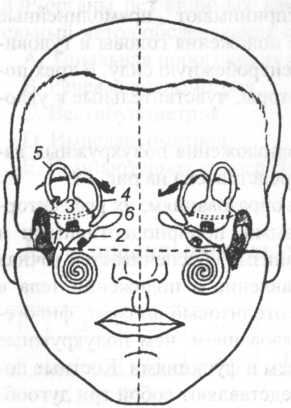
Рис. 18. Анатомо-топографическое расположение полукружных каналов: 1 — столбик; 2 — мешочек; 3 — боковой (горизонтальный) полукружный канал; 4 — задний (са- гиттальний) полукружный канал; 5 — передний (фронтальный) полукружный канал; 6 — эндолимфатический мешочек
Эти отделы отличаются строением и функциями. Костные полукружные каналы по конфигурации представляют собой три дугообразные трубки, расположенные во взаимно перпендикулярных плоскостях — горизонтальной, сагиттальной и фронтальной. Полости костных полукружных каналов и преддверия заполнены прозрачной жидкостью — перилимфой. В перилимфе в подвешенном состоянии расположен перепончатый лабиринт: перепончатые полукружные протоки, маточка и мешочек преддверия, повторяющие форму костного лабиринта. Полость перепончатого лабиринта заполнена эндолимфой, которая по биохимическому составу отличается от перилимфы. Каждый полукружный канал на одном конце имеет расширение (ампулу) с небольшим выступом, или гребешком (crista ampullaris), на котором расположены чувствительные нейроэпителиальные клетки (рис. 19). Внутренняя поверхность перепончатого лабиринта выстлана эпителием, в ампулах его содержатся клетки двух типов: на верхушке ампулярного гребешка — рецепторные нейроэпителиальные, возле основания и между ними — опорные (поддерживающие). Нейроэпителиальные клетки заканчиваются чувствительными волосками, образующими своеобразную кисточку (cupula), или заслонку (valvula) (см. вклейку, рис. 20). Заслонка свободно размещается в отверстии перепончатого протока, имеет малую массу и способна легко отклоняться в ту или иную сторону при незначительном перемещении эндолимфы. Такое смещение чувствительных волосинок (ресничек) вызывает раздражение нейроэпителиальных клеток и вестибулярного нерва. В волосковом аппарате различают киноцилии и стереоцилии. Киноцилии более длинные и подвижные, чем стереоцилии.
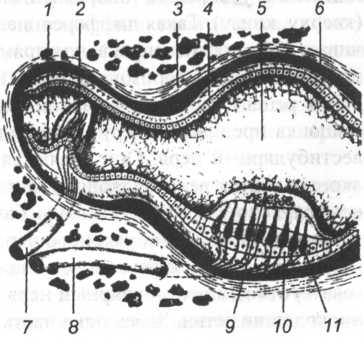
Рис. 19. Схема строения ампулярного гребня и столбика (по А.Х. Миньковскому, 1974): 1 — ампулярний гребень; 2 — кисточка (cupula); 3 — костный лабиринт; 4 — перепончатый лабиринт; 5 — полость столбика; 6 — перилимфати- ческое пространство; 7—ампульний нерв; 8 — маточковый нерв; 9 — статоконии (отолиты); 10 — опорные клетки; 11 — нейроэпителиальные клетки.
При механическом смещении ресничек волосковых клеток изменяется электрический заряд в эндолимфе и наступает соответственно возбуждение или торможение активности рецепторной клетки. Перемещение фибрилл (ресничек) волоскового аппарата от стереоцилии до киноцилии сопровождается отрицательным потенциалом (деполяризацией) в эндолимфе, приводит к возбуждению рецепторных клеток и усилению афферентных импульсов. И наоборот, смещение ресничек от киноцилии в сторону стереоцилии сопровождается положительным потенциалом (гиперполяризацией), что тормозит активность рецепторных клеток.
В перепончатых маточке и мешочке преддверия содержатся рецепторные образования — пятна (macula utriculi et sacculi). Они представлены нейросенсорными (рецепторными) и опорными клетками. Рецепторные клетки имеют короткие волоски, опорные — длинные. Длинные волоски опорных клеток тесно переплетаются между собой, образуя что-то наподобие сетки. В ее петлях, содержащих студневидную массу, расположены очень мелкие кристаллы кальция фосфата и кальция карбоната — статоконии. Все это образует мембрану статоко- ний (см. вклейку, рис. 21). При возникновении прямолинейного ускорения мембрана статоконий скользит, оказывая давление на волокна чувствительных нейросенсорных клеток, что приводит к раздражению вестибулярного нерва. Статоконии маточки расположены сагиттально, статоконии мешочка — горизонтально. Первые реагируют преимущественно на прямолинейные ускорения, направленные в сагиттальной плоскости относительно тела человека, а вторые — на прямолинейные ускорения, направленные горизонтально и вертикально (кверху, книзу). Такая дифференциация отолитового аппарата обеспечивает восприятие направления прямолинейного движения и контроль положения тела в горизонтальной и вертикальной плоскостях.
От рецепторных образований полукружных протоков, маточки и мешочка преддверия берут начало нервные волокна, формирующие вестибулярный нерв. Они являются дистальными отростками биполярных клеток вестибулярного ганглия, расположенного во внутреннем слуховом проходе. Аксоны биполярных клеток образуют центральную часть вестибулярного нерва, который входит в ствол мозга на уровне мостомозжечкового угла. В ромбовидной области продолговатого мозга вестибулярный нерв подразделяется на восходящие и нисходящие ветви. Здесь одна часть нисходящих волокон заканчивается в нижнем ядре, другая — в медиальном и латеральном ядрах, восходящие волокна заканчиваются в верхнем ядре.
Дальнейший путь вестибулярного нерва и его локализация в коре большого мозга еще недостаточно изучены. Это положение можно объяснить сложностью вестибулярного ядерного комплекса по своей структуре и наличием его широких афферентно-эфферентных связей с различными анализаторами и системами организма. Латеральное вестибулярное ядро, в котором прерывается вестибулярный нерв, функционально связано со спинным мозгом. Здесь берет начало вестибулоспинальный путь (tractus vestibulospinalis), который заканчивается в двигательных клетках передних отрогов спинного мозга и предопределяет анимальные реакции в виде изменения тонуса мышц конечностей и туловища. В верхнем ядре начинается восходящая система волокон к среднему мозгу, высшим его уровням, которые частично заканчиваются в глазодвигательных ядрах, зрительном бугре и обеспечивают окуломоторные реакции (нистагм глаз). Медиальное вестибулярное ядро тесно связано с мозжечком, спинным и средним мозгом, с помощью которых осуществляется влияние вестибулярного аппарата на организм.
Ориентировочная схема проводящих путей и связей вестибулярного анализатора с ядрами мозжечка, глазодвигательными нервами, четверохолмием телом, вегетативными центрами, расположенными в продолговатом мозге, представлена на рис. 22.
Таким образом, вестибулярные ядра обладают широкими анатомическими и функциональными связями с разными структурами мозга и системами организма. Благодаря этому вестибулярный аппарат может оказывать рефлекторное влияние на сенсорные, соматические и вегетативные функции. Однако вестибулярный аппарат имеет не только прямые (афферентные) связи с ЦНС и другими органами, есть и обратные (эфферентные) пути к нему от этих органов, осуществляемые, по мнению большинства исследователей, посредством ретикулярной формации мозга.
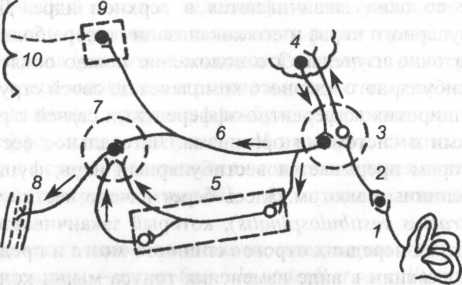
Рис. 22. Общая схема путей и взаимосвязей вестибулярного анализатора (В.Ф. Ундриц, 1960): 1 — лабиринт; 2 — предцверный узел; 3 — преддверные ядра в продолговатом мозге; 4 — ядра мозжечка; 5 — ретикулярная формация; 6 — афферентный путь к ядрам глазодвигательных нервов (7) и зрительного бугра (8); 9 — мышцы глаз, обеспечивающие вестибулярный нистагм; 10 — корковая зона вестибулярного анализатора (височно-теменная область)
Наличие двусторонней связи вестибулярного аппарата с ретикулярной формацией ствола мозга и другими системами многое объясняет в механизме их взаимного влияния, возникновении первичных и вторичных проявлений вестибулярной дисфункции. Данные клиникоэкспериментальных исследований свидетельствуют о том, что раздражение вестибулярного аппарата сопровождается изменениями функции обоняния, зрения, слуха и т. п. С другой стороны, ароматные вещества, а также зрительные, обонятельные, проприоцептивные раздражения вызывают нарушение вестибулярной функции. Поэтому в клинике часто наблюдаются вестибулярные расстройства у больных с сердечно-сосудистой патологией, черепно-мозговыми травмами, заболеваниями внутренних органов. Распознавание первичных и вторичных признаков вестибулярной дисфункции в таком случае имеет большое значение для определения тактики лечения.
Кровоснабжение периферического отдела вестибулярного аппарата осуществляется лабиринтной артерией (a.labyrinthi), которая берет начало от передней и частично средней мозжечковой артерии. Во внутреннем слуховом проходе лабиринтная артерия разделяется на преддверную {a. vestibularis) и улитковую (a. cochlearis). Первая обеспечивает кровью преддверие, вторая — улитку.
Иннервируется лабиринт VIII парой черепных нервов (n. vestibulo- cochlearis). Нерв выходит из продолговатого мозга, входит вместе с лицевым нервом во внутренний слуховой проход и здесь разделяется на два корешка — преддверный (radix vestibularis) и улитковый (radix cochlearis).
Преддверный корешок образует во внутреннем слуховом проходе большой узел (gangl. vestibularis), который делится на верхнюю и нижнюю части и образует ряд нервных ветвей (п. utricularis, п. saccularis, п. ampullaris),направленных к рецепторным образованиям ампул полукружных каналов, маточки и мешочка преддверия.
Дата добавления: 2016-05-25; просмотров: 3591;