ИНТЕНСИВНОСТЬ СТИМУЛА ХАРАКТЕРИЗУЕТСЯ ПОРОГОМ ОЩУЩЕНИЯ (ВОСПРИЯТИЯ).
Абсолютный порог ощущения — это минимальная интенсивность стимула, которая создает соответствующее чувство. Дифференциальный порог — это минимальное различие интенсивностей, которое воспринимается субъектом. Это означает, что анализаторы способны дать количественную оценку прироста ощущения в сторону его увеличения или уменьшения.
Пространственные характеристикидействующих стимулов, необходимые для их различения, зависят от: 1)специфических особенностей каждой сенсорной системы и 2)величины рецептивных полей. Например, прикосновение к коже дистальной фаланги пальца руки двух ножек циркуля с расстоянием между ними 2 мм ощущается раздельно, но чтобы ощутить раздельное прикосновение к коже спины, ножки циркуля необходимо раздвинуть до 60 мм. Две точки зрительного поля не сливаются в одну, если отражаемые ими световые лучи попадут на разные рецептивные поля сетчатки. Имеет значение и степень контраста между действующим стимулом и его фоном: хорошо контрастируемые объекты (например, черное на белом) различаются легче, чем мало контрастируемые (черное на сером). Временная характеристикавосприятия действующих стимулов у человека имеет абсолютный порог различения коротких временных отрезков, который соответствует примерно 1/18секунды. Например, 18 зрительных изображений, предъявленных в течение 1 секунды, сливаются в непрерывное движение, 18 прикосновений к коже за 1 секунду воспринимаются как одно, а 18 звуковых колебаний в секунду воспринимаются как один очень низкий звук. Разрешающая способность сенсорных систем для восприятия действующих через малые промежутки времени стимулов ограничена рефрактерным периодом, во время которого система не способна реагировать на предъявленный стимул.
КЛАССИФИКАЦИЯ РЕЦЕПТОРОВ
В основу классификации рецепторов положены следующие принципы:
1. Среда, в которой рецепторы воспринимают информацию (экстеро-, интеро-, проприо- и другие рецепторы).
2. Природа адекватного раздражителя (механо-, термо-, фото-и другие рецепторы).
3. Характер ощущения после контакта с рецепторами (тепловые, холодовые, болевые и др.).
4. Способность воспринимать раздражитель, находящийся на расстоянии от рецептора — дистантный (обонятельный, зрительный) или при непосредственном контакте с ним — контактный (вкусовой, тактильный).
5. По количеству воспринимаемых модальностей (раздражителей) рецепторы могут быть мономодальными (например, световой) и полимодальными (механический и температурный).
6. Морфологические особенности и механизмы возникновения возбуждения. Различают первичночувствующие (обонятельные, тактильные) и вторичночувствующие рецепторы (зрения, слуха, вкуса).
Этапы деятельности анализаторной системы
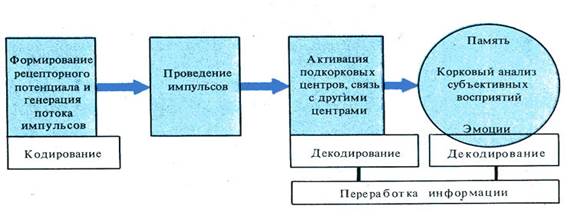
Кодирование информации в рецепторах
Кодирование качества осуществляется за счет избирательной чувствительности рецептора к адекватному с низким порогом возбуждения раздражителю, т.е. рецептор «узнает» свой стимул (глаз - свет, ухо - звук) и за счет существования цепей модально-специфичных нейронов, соединенных синапсами в определенную жесткую цепь, передающую информацию только от своего рецептивного поля. Интенсивность или сила стимула кодируется увеличением частоты ПД, которая, в свою очередь, зависит от величины рецепторного потенциала. Пространственное кодирование.Каждое рецептивное поле имеет свое представительство в определенных структурах центральной нервной системы. Кроме того, рецептивные поля перекрываются , что обеспечивает надежность в работе системы и позволяет слабым раздражителям вступать в контакт с наиболее чувствительными рецепторами и вовлекать в возбуждение менее чувствительные. Кодирование во времени происходит за счет изменения частоты импульсов и продолжительности межимпульсных интервалов.
ФИЗИОЛОГИЯ ЗРИТЕЛЬНОГО АНАЛИЗАТОРА
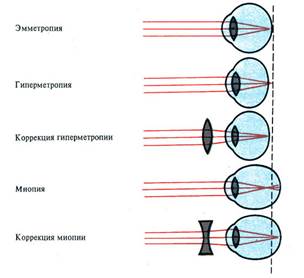
Прежде чем попасть на сетчатку, световые лучи последовательно проходят через роговицу, жидкость передней камеры глаза, хрусталик и стекловидное тело, вместе образующие оптическую систему глаза. На каждом из этапов этого пути свет преломляется и в результате на сетчатке возникает уменьшенное и перевернутое изображение наблюдаемого предмета, этот процесс называется рефракцией.
Особенность топографии палочек и колбочек состоит в том, что они обращены своими наружными светочувствительными сегментами к слою пигментных клеток, т.е. в сторону, противоположную свету. Палочки более чувствительны к свету, чем колбочки. Так, палочку может возбудить всего один квант света, а колбочку — больше сотни квантов. При ярком дневном свете максимальной чувствительностью обладают колбочки, которые сконцентрированы в области желтого пятна или центральной ямки. При слабом освещении в сумерках наиболее чувствительна к свету периферия сетчатки, где находятся в основном палочки. При действии кванта света в рецепторах сетчатки происходит цепь фотохимических реакций, связанных с распадом зрительных пигментов родопсина и йодопсина и их ресинтез в темноте.
Родопсин- пигмент палочек-высокомолекулярное соединение, состоящее из ретиналя — альдегида витамина А и белка опсина. При поглощении кванта света молекулой родопсина 11 - цис-ретиналь выпрямляется и превращается в транс-ретиналь. Это происходит в течение 1-12ceк. Белковая часть молекулы обесцвечивается и переходит в состояние метародопсина II, который взаимодействует с примембранным белком трансдуцином. Последний запускает реакцию обмена гуанозиндифосфата (ГДФ) на гуанозинтрифосфат (ГТФ), чтоприводит к усилению светового сигнала. ГТФ вместе с трансдуцином активирует молекулу примембранного белка — фермента фосфодиэстеразы, который разрушает молекулу циклического гуанозинмонофосфата (цГМФ), вызывая еще большее усиление светового сигнала. Падает содержание цГМФ и закрываются каналы для Na+ и Са2+, что приводит к гиперполяризации мембраны фоторецептора и возникновению рецепторного потенциала. Возникновение гиперполяризации на мембране фоторецептора отличает его от других рецепторов, например слуховых, вестибулярных, где возбуждение связано с деполяризацией мембраны. Гиперполяризационный рецепторный потенциал возникает на мембране наружного сегмента, далее распространяется вдоль клетки до ее пресинаптического окончания и приводит к уменьшению скорости выделения медиатора - глутамата. Для того чтобы рецепторная клетка могла ответить на следующий световой сигнал, необходим ресинтез родопсина, который происходит в темноте (темновая адаптация) из цис-изомера витамина А, поэтому при недостатке в организме витамина А развивается недостаточность сумеречного зрения («куриная слепота»).
Фоторецепторы сетчатки связаны с биполярной клеткой с помощью синапса. При действии света уменьшение глутамата в пресинаптическом окончании фоторецептора приводит к гиперполяризации постсинаптической мембраны биполярной нервной клетки, которая также синаптически связана с ганглиозными клетками. В этих синапсах выделяется ацетилхолин, вызывающий деполяризацию постсинаптической мембраны ганглиозной клетки. В аксональном холмике этой клетки возникает потенциал действия. Аксоны ганглиозных клеток образуют волокна зрительного нерва, по которым в мозг устремляются электрические импульсы.
Чтобы фокусировать на сетчатке световые лучи, отраженные от близко расположенных предметов, оптическая система глаза должна преломлять их тем сильней, чем ближе расположен наблюдаемый объект. Механизм, с помощью которого глаз настраивается на рассмотрение удаленных или близких предметов и в обоих случаях фокусирует их изображение на сетчатку, называется аккомодацией.
Гладкие мышцы ресничного тела, управляемые парасимпатическими нейронами, регулируют натяжение цинновой связки: при полном расслаблении мышц связка натягивает капсулу хрусталика, заставляя его принимать максимально уплощенную форму, необходимую для рассмотрения далеких предметов.
МЕХАНИЗМ АККОМОДАЦИИ ГЛАЗА
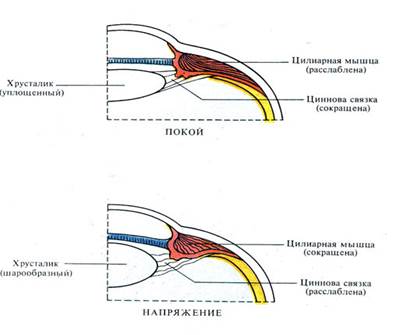
СХЕМА ХОДА ЛУЧЕЙ ЧЕРЕЗ ПРЕЛОМЛЯЮЩИЕ СРЕДЫ ГЛАЗА
Движения глаз. При наблюдении за движущимися в зрительном поле объектами, а также при движении человека относительно окружающего мира происходят следящие движения глаз,благодаря которым изображение в одной и той же области сетчатки сохраняется неизменным. При зрительном восприятии неподвижных объектов, имеющих многочисленные детали формы, а также во время чтения происходят быстрые движения глаз, предназначенные для фиксации наиболее информативных деталей объекта. Изображения глаз проецирует не вообще на сетчатку, а на ту ее область, которая обладает максимальной разрешающей способностью. Это центральная ямка,представляющая собой небольшое углубление диаметром около 3 мм в центре сетчатки.
При рассматривании любых объектов глаза ежесекундно совершают около трех очень быстрых непроизвольных и субъективно не ощущаемых движений, которые называются саккадами. Благодаря таким движениям изображение на сетчатке регулярно смещается, вызывая раздражение разных фоторецепторов. Необходимость саккад объясняется свойством зрительной системы сильнее реагировать на изменяющееся раздражение (появление или исчезновение стимула), тогда как на постоянную стимуляцию она отвечает слабо.
РЕЦЕПТИВНЫЕ ПОЛЯ КЛЕТОК СЕТЧАТКИ
Существуют два пути для передачи сигналов от фоторецепторов к ганглиозной клетке:
1. Прямой путьначинается от фоторецепторов, расположенных в центре рецептивного поля и образующих синапс с биполярной клеткой, которая через другой синапс действует на ганглиозную клетку.
2. Непрямой путьберет начало от фоторецепторов периферии рецептивного поля, которая с центром состоит в реципрокных отношениях, обусловленных тормозным действием горизонтальных и амакриновых клеток (латеральное торможение).
Примерно половина ганглиозных клеток возбуждается действием света на центр рецептивного поля и тормозится при действии светового стимула на периферию рецептивного поля. Такие клетки принято называть оn-нейронами.
Другая половина ганглиозных клеток возбуждается действием светового раздражителя на периферию рецептивного поля и тормозится в ответ на световую стимуляцию центра рецептивного поля — они получили название оff-нейронов.
Рецептивные поля ганглиозных клеток обоих типов в сетчатке представлены поровну, чередуясь друг с другом. Оба типа клеток очень слабо отвечают на равномерную диффузную засветку всего рецептивного поля, а наиболее сильным раздражителем для них является световой контраст, т. е. различная интенсивность засветки центра и периферии. Именно контрастирование деталей изображения дает необходимую информацию для зрительного восприятия в целом, тогда как абсолютная интенсивность отраженного от наблюдаемого объекта света не столь важна. Восприятие граней, т. е. восприятие контраста между соседними поверхностями с разной освещенностью, является наиболее информативным признаком изображения, определяющим протяженность и позиции разных объектов.
Дата добавления: 2016-05-25; просмотров: 1161;