Взаимодействие аллельных генов
Взаимодействие аллельных генов называют также внутриаллельным.
Выделяют следующие его виды:
- полное доминирование,
- неполное доминирование,
- сверхдоминирование,
- кодоминирование,
- межаллельная комплементация,
- аллельное исключение.
Ранее мы рассмотрели полное доминирование, описанное Менделем. Известно также неполное доминирование.
При неполном доминированиигетерозигота имеет фенотип, промежуточный между фенотипами гомозигот. Например, у растения ночная красавица (Mirabilis jalapa) имеются гомозиготные линии с красными (АА) и белыми (аа) цветками. Гибриды F1 являются гетерозиготами Аа и имеют цветки розовой окраски. Во втором поколении наблюдается расщепление 1:2:1 как по фенотипу, так и по генотипу. В целом, 1 и 2 законы Менделя соблюдаются с поправкой на промежуточный фенотип у гетерозигот.
Другие примеры неполного доминирования: окраска цветков у львиного зева, окраска оперения у андалузских кур, шерсти у коров (красная и белая; промежуточная – чалая масть).
Причина промежуточного проявления признака у гетерозигот: аллель А находится в генотипе гетерозиготы в единственном числе, поэтому проявляется слабее, чем в геноме гомозиготы АА (двойная доза генов, а значит, и продуктов генов!). Для неполного доминирования важным показателем является экспрессивность – степень выраженности признака, зависящая от дозы генов (аллелей) и от условий окружающей среды (влияние УФ – света на окраску кожи).
Экспрессивность снижается в ряду: АА> Аа > аа.
Примеры неполного доминирования генов у людей: гомозиготы по аллелю серповидноклеточности эритроцитов (HвS) не переживают детский возраст из-за анемии и сопутствующих расстройств, гетерозиготы почти здоровые люди, но при подъеме на высоту они испытывают кислородное голодание. Т. е. в данном случае речь идет о неполном доминировании гена нормального гемоглобина (НвА). Подобное соотношение наблюдается и при других наследственных заболеваниях человека, вызванных мутантными рецессивными аллелями.
При сверхдоминировании доминантный ген в гетерозиготном состоянии проявляет себя сильнее, чем в гомозиготном. У дрозофилы, например, имеется летальный рецессивный ген плодовитости (а). Мухи, гомозиготные (АА) по доминантному гену плодовитости А имеют нормальную жизнеспособность, а гетерозиготы (Аа) живут дольше и более плодовиты, чем доминантные гомозиготы.
Кодоминирование – это одновременное проявление в фенотипе обоих аллелей.
Примеры: поверхностные белки эритроцитов, определяющие группы крови системы АВО у людей; различные формы ферментов с неодинаковой электрофоретической подвижностью.
Группы крови: 3 аллеля: I0, IA и IB
1я: I0 I0
2я: IА IА или IА I0
3я: IВ IВ или IВ I0
4я: IА IВ
В гетерозиготах IА I0 и IВ I0 наблюдается полное доминирование аллелей IA и IB над I0, а в гетерозиготе IА IВ проявляются оба аллеля, вызывая образование антител против обоих антигенов.
Другой пример: образование в гетерозиготном организме двух форм одного и того же белка с различной электрофоретической подвижностью.
Межаллельная (внутриаллельная) комплементация относится к достаточно редко встречаемым способам взаимодействия аллельных генов. В этом случае возможно формирование нормального признака D у организма, гетерозиготного по двум мутантным аллелям гена D(D'D''). Допустим, что ген D отвечает за синтез какого-то белка, который имеет четвертичную структуру, состоящую из нескольких одинаковых пептидных цепей. Мутантный аллель D' определяет синтез измененного пептида D', а мутантный аллель D'' приводит к синтезу другой, но тоже измененной структуры пептида D''. Можно представить ситуацию, когда взаимодействие таких измененных пептидов (D' и D'') при формировании четвертичной структуры, как бы взаимно компенсируя эти изменения, обеспечивает образование белка с нормальными свойствами. В то же время отдельно взаимодействующие пептиды D' или D'' формируют аномальные белки. Таким образом, с определенной вероятностью у гетерозигот D'D'' в результате межаллельной комплементации может формироваться нормальный признак за счет белка с нормальными свойствами.
Аллельное исключение – такой вид взаимодействия аллельных генов, который связан с невозможностью присутствия в одной клетке двух активно работающих аллелей одного гена.
Пример: в клетках организма женщины случайным образом происходит инактивация одной из Х – хромосом (тельце Барра).
Т. е. организм женщины мозаичен по функционирующей Х - хромосоме со всеми ее аллелями. Поэтому любой мутантный рецессивный аллель, если он присутствует в Х – хромосоме, будет проявляться в отдельных участках тела женщины, например, отсутствие потовых желез и др.
Таким образом, даже процесс формирования элементарного признака – синтез полипептида - зависит от взаимодействия, по меньшей мере, двух аллельных генов, и конкретный результат определяется конкретным сочетанием их в генотипе.
5. Дигибридное скрещивание. 3й закон Менделя
Изучив наследование одной пары аллелей, Мендель решил проследить наследование двух признаков одновременно. Для этой цели он использовал гомозиготные растения гороха, отличающиеся по двум парам альтернативных признаков: семена желтые гладкие и зеленые морщинистые. Такое скрещивание, при котором родители различаются по двум парам альтернативных признаков, называется дигибридным. Если изучаются 3 и более пар признаков, то скрещивание полигибридное.
В результате такого скрещивания в первом поколении Мендель получил растения с желтыми гладкими семенами. Этот результат показал, что закон единообразия гибридов первого поколения проявляется не только при моногибридном, но и при полигибридном скрещивании, если родительские формы гомозиготны.
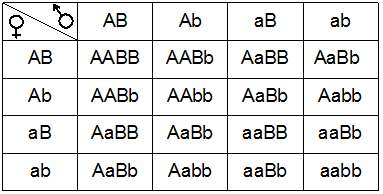
Затем Мендель скрестил гибриды первого поколения между собой. Для анализа результата этого скрещивания используем решетку Пеннета.
В результате свободного комбинирования 4 типов гамет в зиготы попадают гены во всех возможных 16ти комбинациях. В потомстве выявляются 4 фенотипических класса: примерно 9 частей растений с горошинами желтыми гладкими (А-В-), 3 части – с желтыми морщинистыми (А-вв), 3 части – с зелеными гладкими (ааВ-), 1 часть – с зелеными морщинистыми (аавв), т. е. расщепление примерно 9:3:3:1. Два фенотипических класса из четырех – абсолютно новые, отличные от родительских форм. При этом количество генотипических классов равно 9.

 P: AABB × aabb
P: AABB × aabb
 G: AB ab
G: AB ab
F1: AaBb

 P(F1): AaBb × AaBb
P(F1): AaBb × AaBb
Если проанализировать расщепление отдельно по каждой из пар альтернативных признаков (желтый и зеленый цвет, гладкая и морщинистая поверхность), то получится: ~3:1→ желтые : зеленые и
~ 3:1→ гладкие : морщинистые,
т. е. каждая пара признаков дала расщепление в F2 независимо от другой пары. Это явилось результатом случайного комбинирования генов при образовании зигот, что и привело к образованию двух новых фенотипических классов, отличных от родительских форм.
Отсюда вытекает третий закон Менделя – закон независимого комбинирования признаков: при дигибридном скрещивании гетерозиготных организмов у гибридов наблюдается независимое комбинирование признаков и генов разных аллельных пар.
Для объяснения результатов скрещивания, проведенного Г. Менделем, Уильям Бэтсон (1902 г.) предложил гипотезу «чистоты гамет». Ее можно свести к следующим двум основным положениям:
1) из каждой пары аллелей в гамету попадает только один ген;
2) у гибридного организма гены не смешиваются, а находятся в чистом аллельном состоянии.
Эта гипотеза после открытия механизмов мейоза стала очевидным фактом и называется теперь правилом «чистоты гамет».
В общем случае для полигибридного скрещивания справедливы формулы, которые показаны в таблице:
Таблица – Количество классов потомков в F2 при полигибридном скрещивании
| Число пар генов | Число различных типов га-мет, образуемых гибридом F1 | Количество в F2… | ||
| комбинаций аллелей | фенотипических классов | генотипических классов | ||
| … | … | … | … | … |
| n | 2n | 4n | 2n | 3n |
Дата добавления: 2017-02-04; просмотров: 4176;