Особенности строения и созревания и-РНК
И-РНК у эукариот моноцистронная, т. е. кодирует синтез одного полипептида (как правило), а у прокариот – полицистронная, т. е. определяет синтез сразу нескольких пептидов, например, это – каскад ферментов.
В молекуле и-РНК как прокариот, так и эукариот имеются кодирующие участки и дополнительные последовательности. Перед стартовым кодоном (АУГ или ГУГ) на 5' – конце это – лидерный участок. На 3' – конце после стоп – кодона расположен трейлер. Функция лидерных участков – связывание и-РНК с рибосомой (через комплементарное взаимодействие с рРНК), а функция трейлеров – повышение стабильности молекулы и-РНК.
В полицистронной РНК прокариот между кодирующими участками имеются небольшие межцистронные области. Эти и – РНК сразу после синтеза готовы к трансляции и находятся в цитоплазме.
В отличие от них, молекулы и – РНК эукариот прерывисты и содержат экзоны(кодирующие участки) и интроны(некодирующие).
Сразу после синтеза первичные транскрипты молекулы и – РНК крупные, незрелые, они образуют гетерогенную ядерную РНК (гя РНК), которая долго ждет своего часа. Перед выходом из ядра она подвергается созреванию, или процессингу, в ходе которого происходит модификация концевых участков первичного транскрипта и сплайсинг. Удаление интронов с последующим соединением экзонов называется сплайсингом.
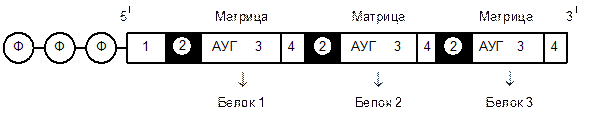
Рис. 1 – Полицистронная матричная РНК прокариот: 1 – лидерный участок, 2 – межцистронные области, 3 – кодирующие области, 4 – трейлер
Модифицирование первичного транскрипта эукариот включает образование колпачка – кэпа, который блокирует 5'-конец и-РНК путем присоединения к его первому нуклеотиду трифосфонуклеозида, содержащего гуанин, связью 5' – 5'. В результате образуется последовательность, в которой остаток гуанина находится в обратной ориентации по отношению к другим нуклеотидам и-РНК. Модификация 5'-конца и-РНК предполагает также метилирование присоединенного гуанина и первых двух – трех оснований первичного транскрипта. Кэпы обеспечивают узнавание молекул и-РНК малыми субъединицами рибосом.
После завершения транскрипции происходит удаление части нуклеотидов на 3'-конце первичного транскрипта и присоединение к нему последовательности, состоящей из 100-200 остатков адениловой кислоты (полиА).
После выхода и-РНК в цитоплазму ее полиА-последовательность постепенно укорачивается под действием ферментов, отщепляющих нуклеотиды на 3'-конце. Возможно, добавление полиА-последовательности в ходе процессинга повышает стабильность и-РНК.
Кроме метилирования в области кэпа в молекуле и-РНК высших эукариот происходит метилирование небольшой части внутренних нуклеотидов с частотой приблизительно одно на тысячу оснований и-РНК.
Созревание и-РНК у эукариот предполагает также удаление из первичных транскриптов интронов, т. е. сплайсинг. Размер интронных участков варьирует от 100 до 10000 нуклеотидов. На долю интронов приходится около 80% всей гяРНК.
В настоящее время описано несколько механизмов сплайсинга. Точность этого процесса достигается действием ферментов, узнающих концевые участки интронов и катализирующих разрыв фосфодиэфирных связей на границе экзон – интрон, а затем образование связей между двумя экзонами.
Установлено активное участие в сплайсинге особых, малых ядерных РНК (мяРНК), образующих комплексы с протеинами (мяРНП). Очевидно, мяРНК своими нуклеотидными последовательностями комплементарно взаимодействуют с концевыми участками интронов, которые образуют при этом замкнутые петли. Расщепление РНК в устье интронной петли приводит к удалению неинформативной последовательности и соединению (сплайсингу) сближенных концов экзонов.
Возможно также, что РНК-транскрипт обладает автокаталитическойспособностью к сплайсингу.
Благодаря преобразованиям в ходе процессинга, зрелые и-РНК эукариот характеризуются большей стабильностью по сравнению с прокариотическими и-РНК, что крайне необходимо для их сохранности в течение длительного времени.
По завершении процессинга в цитоплазму попадает всего 5% гяРНК. Остальная часть расщепляется, не покидая ядра.
Дата добавления: 2017-02-04; просмотров: 1527;