Конвульсанты, действующие на холинореактивные синапсы
Синаптические структуры, в которых медиаторную функцию выполняет ацетилхолин, обнаружены как в центральной нервной системе, так и на периферии.
Процесс передачи сигнала в холинэргических синапсах может быть представлен несколькими этапами:
- синтез медиатора из холина и ацетата при участии энзима холин-ацетил трансферазы (ХАТ) и его депонирование в пресинаптических везикулах;
- выделение ацетилхолина в синаптическую щель (спонтанно и при поступлении в нервное окончание возбуждающего сигнала);
- взаимодействие ацетилхолина с холинорецепторами постсинаптической (по некоторым данным, и пресинаптической) мембраны и инициация стимула в иннервируемой клетке;
- разрушение выделившегося в синаптическую щель нейромедиатора энзимом ацетилхолинэстеразой (АХЭ) и прекращение процесса передачи сигнала;
- захват пресинаптическими структурами высвободившегося холина.
Наиболее уязвимыми для действия высокотоксичных веществ являются этапы выделения ацетилхолина в синаптическую щель, взаимодействия медиатора с холинорецепторами и разрушения его энзимом АХЭ (рисунок 44).
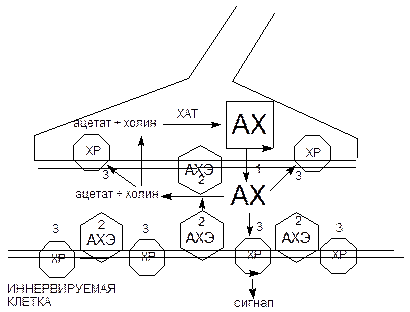
Рисунок 44. Схема функционирования холинэргического синапса
ХР - холинорецептор; АХЭ – ацетилхолинэстераза;
1 - 3 - способы воздействия токсикантов на синапс: 1 - ботулотоксин; 2 - ингибиторы АХЭ; 3 - холинэргические и антихолинэргические средства
Установлено, что нейромедитор и в ЦНС и в ПНС взаимодействует с двумя видами рецепторов: мускариновыми и никотиновыми, отличающимися между собой способностью отвечать на воздействие ряда агонистов (М-холинорецепторы избирательно возбуждаются мускарином; Н-холинорецепторы - никотином), механизмом рецепции сигнала и его передачи на эффекторную клетку (нейрон, миоцит, железистую клетку). Н-холинорецепторы непосредственно связаны с ионными каналами постсинаптической мембраны для Na+, K+, Ca2+. М-холинорецепторы представляют собой структуры, которые также влияют на проницаемость ионных каналов для Na+ и K+. Однако это влияние опосредовано изменением активности сопряженных с рецептором ферментов, регулирующих содержание в иннервируемой клетке вторичных мессенджеров восприятия сигнала (цАМФ, цГМФ, фосфоинозитола и т.д.). И в том и в другом случае действие ацетилхолина на рецепторы приводит к усилению проницаемости постсинаптических мембран для ионов натрия, что сопровождается деполяризацией мембраны и ее возбуждением. Различается скорость генерации постсинаптического сигнала: в Н-холинэргических синапсах изменение конформации ионных каналов развивается практически мгновенно, а в М-холинэргических - постепенно.
Установлено, что в синапсах на пресинаптической мембране локализуются холинорецепторы, выполняющие функцию регуляторов проведения импульса.
Прерывание нервного импульса осуществляется путем гидролиза ацетилхолина ферментом ацетилхолинэстеразой. Механизм взаимодействия ацетилхолина с холинэстеразой изучен достаточно подробно. Известно, что в состав молекулы ацетилхолина входят реакционноспособные атомы, которые образуют две группы: катионную (четвертичный атом азота с тремя метильными радикалами) и сложноэфирную (эфирный кислород и карбонильные углерод и кислород) (рис. 45).
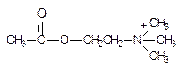
Рисунок 45. Структура ацетилхолина
Соответственно реакционным группам ацетилхолина, в активных центрах холинэстеразы также выделяются два участка: анионный и эстеразный. Предполагается, что анионный участок образован карбоксил-анионом двухосновной аминокислоты. На расстоянии 0,4-0,5 нм от анионного центра расположен эстеразный участок, в котором функциональную роль играет гидроксильная группа аминокислоты серина.
Взаимодействие ацетилхолина с холинэстеразой начинается с ориентации катионной “головки” ацетилхолина на анионном участке фермента. Это приводит к фиксации молекулы ацетилхолина на определенном расстоянии от эстеразного участка, на котором и проходит ферментативный гидролиз медиатора (разрушение эфирной связи между холином и ацетатом).
Для его осуществления гидроксильная группа серина проводит нуклеофильную атаку на соответствующий электрофильный атом субстрата. В результате реакции образуются ацетилированный по серину фермент и холин, сорбированный на анионном участке. В последующем холин отщепляется от анионного участка фермента, а затем происходит и деацетилирование холинэстеразы - восстанавливается ее первоначальная структура.
В ЦНС плотность М-холинэргических синапсов между нейронами существенно выше, чем Н-холинэргических. Наивысшая - в хвостатом, прилежащем, чичевичном ядрах, гиппокампе, гипоталамусе, коре головного мозга, мозжечке млекопитающих и человека.
На периферии М-холинэргические синапсы связывают окончания постганглионарных нервных волокон парасимпатической системы с иннервируемыми ими гладкомышечными волокнами (кишечник, бронхи, миокард) и клетками экзокринных желез (слюнных, желудочно-кишечного тракта, бронхиальных, потовых).
Периферические Н-холинэргические синапсы выявляются главным образом в области окончаний нервных волокон мотонейронов, иннервирующих поперечно-полосатую мускулатуру, волокон, иннервирующих ганглионарные нейроны парасимпатического и симпатического отделов вегетативной нервной системы, а также в каротидном клубочке, хромаффонинных клетках мозгового вещества надпочечников.
Как указывалось ранее, перевозбуждение центральных холинэргических механизмов может приводить к развитию судорожного синдрома. Наибольшей активностью в этом плане обладают ингибиторы АХЭ, способные проникать через гематоэнцефалический барьер (ФОС, карбаматы). Перевозбуждение периферических холинореактивных структур этими токсикантами сопровождается существенным, порой не совместимым с жизнью, нарушением функций органов и систем.
Дата добавления: 2016-04-14; просмотров: 724;