ВЗАИМОДЕЙСТВИЕ НЕАЛЛЕЛЬНЫХ ГЕНОВ
Если несколько генов определяют одно свойство организма (окраску цветка, длину шерсти и др.), то они взаимодействуют друг с другом. При этом в потомстве дигетерозиготы может наблюдаться необычное расщепление - 9:3:4; 9:7; 9:6:1; 13:3; 12:3:1; 15:1. Генетический анализ показывает, что необычные расщепления по фенотипу в F2 представляют видоизменение общей менделевской формулы 9:3:3:1. Известны случаи взаимодействия трех и большего числа генов с изменением обычных формул расщепления.
Наиболее часто встречаются 3 формы взаимодействия неаллельных генов: кооперация, комплементарность, эпистаз и полимерия.
Комплементарность — явление, когда признак развивается только при взаимном действии двух доминантных неаллельных генов, каждый из которых в отдельности не вызывает развитие признака.
Эпистаз — явление, когда один ген (как доминантный, так и рецессивный) подавляет действие другого (неаллельного) гена (как доминантного, так и рецессивного). Ген-подавитель (супрессор) может быть доминантным (доминантный эпистаз) или рецессивным (рецессивный эпистаз).
Полимерия — явление, когда несколько неаллельных доминантных генов отвечают за сходное воздействие на развитие одного и того же признака. Чем больше таких генов присутствуют в генотипе, тем ярче проявляется признак. Явление полимерии наблюдается при наследовании количественных признаков (цвет кожи, вес тела, удойность коров).
В противоположность полимерии наблюдается такое явление, как плейотропия — множественное действие гена, когда один ген отвечает за развитие нескольких признаков.
Комплементарность. Комплементарными или дополнительными называют такие доминантные гены, которые при совместном нахождении в генотипе (А-В-) обусловливают развитие нового признака по сравнению с действием каждого гена, в отдельности (A-bb или ааВ-).
Расщепление 9:3:3:1.Так, у дрозофилы встречается коричневая и ярко-красная окраска глаз. Обе эти окраски рецессивны к красной окраске (дикий тип). При скрещивании мух с коричневыми и ярко-красными глазами гибриды F1 оказываются красноглазыми, а в F2 наблюдается расщепление на 4 фенотипических класса в отношении 9/16 красные : 3/16- ярко-красные: 3/16 коричневые и 1/16 белые (рис. 2).
Отличие исходных форм по одной паре признаков могло бы свидетельствовать о моногенных различиях между ними. Однако в F1 вместо доминирования одного из признаков появляется новое качество - красная окраска, а в F2 осуществляется дигибридное расщепление с тем лишь отличием от менделевского, что оно идет по одному, а не по двум свойствам (только окраска глаз). При этом здесь проявляется еще один новый признак - белый цвет глаз. Таким образом, генетический анализ свидетельствует о том, что в этом скрещивании участвуют не одна, а две пары генов.
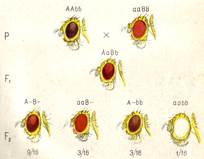
Рисунок 2. Наследование окраски глаз у дрозофилы (комплементарность). Окраска глаз: а —ярко-красная; в —коричневая.
Мы можем сделать вывод, что гены А и В вместе определяют красную окраску глаз дикого типа, а - ярко-красную, в - коричневую. Тогда генотип мух с коричневыми глазами можно обозначить AAbb, с ярко-красными — ааВВ, генотип красноглазых гибридов — АаВb и белоглазых мух aabb. Фенотипические радикалы полученных в F2 классов могут быть представлены как 9 А-В-, 3 ааВ-, 3 A-bb, 1 aabb.
Биохимический анализ глазных пигментов показал, что красная окраска глаз обеспечивается тремя видами пигментов: ярко-красным, коричневым и желтым.
Рецессивный ген а блокирует образование коричневого пигмента, вследствие чего развиваются ярко-красные глаза, другой рецессивный ген — b блокирует одновременно образование красного и желтого пигментов, и поэтому образуется только коричневый пигмент. В F1 объединяются доминантные аллели этих генов, и поэтому синтезируются все пигменты, дающие в совокупности красную окраску глаз. Белоглазые мухи, появляющиеся в F2, являются результатом одновременного блокирования синтеза всех трех пигментов.
Аналогичное наследование встречается и у растений. Например, окраска плодов у томатов (Lycopersicon esculeritum) обусловливается каротиновыми пигментами, имеющими огромное значение в синтезе витаминов. Генетический анализ показывает, что красная окраска плодов определяется взаимодействием комплементарных доминантных генов R и Т, оранжевые плоды образуются на растениях с генотипом R-tt, желтые — с генотипом rrТ-, промежуточные желто-оранжевые — rrtt. Здесь также расщепление в F2 соответствует генетической формуле дигибридного скрещивания 9:3:3: 1.
Таким образом, в случае, когда каждый из двух рецессивных неаллеальных генов проявляет самостоятельный фенотипический эффект, расщепление в F2 по фенотипу соответствует менделевскому отношению 9:3:3:1, так как каждый из четырех классов имеет свой особый фенотип.
Расщепление 9:7. Если же рецессивные аллели дают одинаковый фенотипический эффект, характер расщепления меняется. Например, у белого клевера (Trifolium repens) имеются формы с высоким и низким содержанием цианида. При скрещивании их в F1 доминирует первое свойство, а в F2 наблюдается расщепление, близкое к отношению 3:1. Следовательно, эти альтернативные признаки определяются одной парой аллелей. Но иногда при скрещивании двух растений клевера с низким содержанием цианида гибриды F1 имеют много цианида, а в F2 расщепление оказывается близким к отношению 9/16 с высоким содержанием цианида и 7/16 — с низким.
Чтобы выяснить, укладывается ли это расщепление в схему дигибридного менделевского расщепления, представим, что у каждой исходной расы клевера имеется в гомозиготном состоянии лишь по одной из доминантных аллелей (LLhh или llHH), которые при взаимодействии определяют развитие цианида. Поскольку у гибрида первого поколения F1: присутствуют доминантные аллели обоих генов L-H-, в его листьях будет много цианида. В F2 : происходит расщепление в отношении 9/16 L-H-: 3/16 L-hh : 3/16 llН-: 1/16 llhh. Каждый из доминантных генов самостоятельно не может обусловить выработку большого количества цианида, поэтому у растений с генотипами L-hh и llН — мало цианида, и в F2 наблюдается расщепление по фенотипу в отношении 9 : 7.
Генетический анализ нашел подтверждение в биохимическом анализе. Оказалось, что цианид в листьях клевера образуется из глюкозида линамарина под действием фермента линамаразы. Химический анализ листьев клевера разных генотипов проливает свет на характер взаимодействия этих двух пар генов. Экстракт растений L-H- в норме содержит цианид. Дли того чтобы цианид образовался в листьях растений L-hh, необходимо добавить линамаразу, а в 11Н- линамарин. В растениях же llhh при добавлении любого компонента цианид не образуется. Следовательно, мы можем сделать вывод, что ген L обеспечивает образование линамарина, а ген H вырабатывает фермент линамаразу, превращающий линамарин в цианид. Переход гена L в рецессивное состояние l прерывает реакцию образования линамарина, а ген h блокирует образование фермента. Таким образом, в данном случае совместный генетический и биохимический анализы дают представление о механизме взаимодействия генов (табл. 2).
Таблица 2
Образование цианида экстрактами растений клевера разных генотипов
| Что исследуется Генотип | Только экстракт | Экстракт + Линамарин | Экстракт + линамараза |
| L-H- L-hh 11Н llhh | + | + + | + + |
Подобный тип взаимодействия генов, дающий в F2 расщепление 9:7, найден у многих растений, животных и человека. Так, например, наследуется пурпурная и белая окраска цветка у душистого горошка (Lathyrus odoratus), желтая и белая окраска коконов у шелкопряда, нормальный слух и глухота у человека и т. п.
Расщепление в F2 по фенотипу 9 : 7 есть видоизменение расщепления 9:3:3:1, определяемое тем, что и доминантные и рецессивные гены не имеют самостоятельного фенотипического проявления.
Расщепление 9:3:4. До сих пор были рассмотрены примеры комплементарного взаимодействия, при котором каждый из доминантных генов в отдельности не обладал способностью вызвать развитие признака. Известны, однако, случаи, когда оба доминантных комплементарных гена характеризуются самостоятельным проявлением. В соответствии с этим меняется и характер расщепления в F2. Рассмотрим наследование трех типов окраски шерсти у кроликов (Lepus cuninculus) — дикой рыжевато-серой (агути), черной и белой. Окраска дикого типа зависит от наличия гена, распределяющего пигмент по длине волоса. Каждая шерстинка у кроликов агути имеет посередине желтое кольцо, а в основании и на конце — черный пигмент. Такое зонарное распределение пигментов и создает окраску агути, свойственную всем диким грызунам.
У черных кроликов шерстинки по всей длине окрашены равномерно в черный цвет. Белые кролики с красной радужной оболочкой глаз (альбиносы) вовсе лишены пигмента.
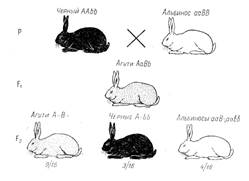
Рисунок 3. Наследование окраски шерсти у кроликов (комплементарность). Окрас шерсти: А – окрашенность; а - альбинизм; В - зонарная (агути); b - черная.
При скрещивании черных кроликов с белыми все гибриды оказываются агути, а в F2 наблюдается расщепление в отношении 9/16 агути: 3/16 черных: 4/16 белых (рис. 3). Если провести анализ этого скрещивания в начале по наличию и отсутствию пигмента, не обращая внимания на его качество, то можно прийти к выводу, что окрашенность доминирует над неокрашенностью, а в F2 наблюдается расщепление на 12 окрашенных (9 + 3) и 4 белых, т. е. 3: 1. В то же время в F2 осуществляется расщепление на 9 агути и 3 черных (3: 1). Гены можно обозначить следующим образом: А - наличие окраски, а - отсутствие ее, В - окраска агути, b - черная. Тогда исходные кролики-альбиносы являются, очевидно, гомозиготными по рецессивному гену отсутствия окраски и доминантному гену агути (ааВВ), а черные кролики - гомозиготными по доминантному гену наличия окраски и рецессивному гену черной окраски (ААbb). У гибридов F1 (АаВb) вследствие взаимодействия доминантных аллелей обоих генов развивается окраска типа агути. Такая же окраска характерна и для 9/16 особей в F2 с генотипом А-В-. Черными в F2 оказываются кролики, имеющие генотип A-bb, а белыми — все остальные (ааВ- и aabb) в силу отсутствия у них гена А, определяющего образование пигмента. Ген В в отсутствии гена А не проявляется.
Подобный тип наследования широко распространен в природе. Например, у ржи (Secale cereale) скрещивание белозерных растений с желтозерными дает в F1 только зеленую окраску зерна, а в F2 расщепление в отношении 9 зеленых: 3 желтых: 4 белых [9:3: (3+1)]. Аналогично наследуется белая, красная и чалая масти у крупного рогатого скота и т. п.
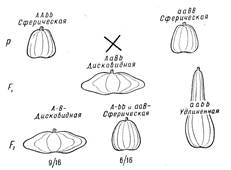
Расщепление 9:6:1. В ряде случаев комплементарные гены, способные к самостоятельному проявлению, при отсутствии дополнительного гена могут давать каждый в отдельности сходный фенотипический эффект. Характер расщепления дигетерозиготы в F2 при этом также изменяется. Так, у тыквы (Cucurbita pepo) имеются сорта с разной формой плода: сферической, дисковидной и удлиненной (рис. 4). Сферическая форма плода является рецессивной но отношению к дисковидной. От скрещивания растений с плодами сферической формы, но имеющих разное происхождение, получаются гибридные растения, дающие дисковидные плоды. В потомстве у этих растений в F2 появляются три фенотипических класса в отношении: 9/16 с дисковидными плодами, 6/16 — со сферическими и 1/16 — с удлиненными. Нетрудно понять, что и здесь имеет место взаимодействие двух генов, определяющих форму плода. Каждый из доминантных комплементарных генов обусловливает развитие плодов сферической формы, а их взаимодействие приводит к образованию дисковидных плодов. Взаимодействие рецессивных аллелей этих генов определяет развитие плодов удлиненной формы. Таким образом, и здесь видоизменяется обычное дигибридное расщепление [9: (3 + 3):1].
Подобный тип взаимодействия наблюдается в наследовании окраски щетины у свиней (Sus scrofa). При скрещивании двух разных пород с песочной окраской в F1 появляется красная окраска, а в F2 расщепление на 9 красных, 6 песочных и 1 белую.
 1/16
1/16
Рисунок 4. Наследование формы плода у тыквы (комплементарность).
Рассматривая примеры комплементарного действия генов, можно убедиться, что оно иногда приводит к развитию у гибридов признаков, несвойственных исходным формам, т. е. к новообразованиям. Зачастую эти «новообразования» являются признаками, свойственными диким предкам данных видов, например окраска агути у кроликов и т. п. У диких предков домашних животных и растений доминантные гены комплементарного действия поддерживались естественным отбором вместе в одном генотипе. При одомашнивании с помощью скрещиваний и искусственного отбора комплементарные гены разобщились. Генотип АаВb разлагался селекционерами на генотипы AAbb и ааВВ. Поэтому при скрещивании и наблюдается иногда как бы возврат к признакам диких предков.
Эпистаз. При доминировании действие одной аллели подавляются другой аллелью этого же гена: А>а, В>b и т. д. Но существует взаимодействие, при котором один ген подавляет действие другого, например А>В или B>A, а>В или b>А и т. д.
Такое явление называется эпистазом. Гены, подавляющие действие других генов, называются супрессорами или ингибиторами. Они могут быть как доминантными, так и рецессивными. Гены-супрессоры известны у животных, растений и микроорганизмов. Обычно они обозначаются I или S.
Эпистаз принято делить на два типа: доминантный и рецессивный.
Под доминантным эпистазом понимают подавление одним доминантным геном действия другого гена.
Расщепление 13:3.Из многих примеров доминантного эпистаза приведем лишь некоторые. Так, у льна (Linura usitatissimum) наряду с формами, имеющими нормальные лепестки, встречаются растения с гофрированными лепестками. При скрещивании двух форм с нормальными лепестками, имеющих разное происхождение, в F1 все гибриды имеют нормальные лепестки, а в F2 получается расщепление: 13/16 растений с нормальными лепестками и 3/16 - с гофрированными. Характер расщепления свидетельствует о том, что форма лепестков определяется двумя парами генов. В таком случае одно из исходных растений должно нести в скрытом состоянии ген гофрированности лепестков, действие которого подавлено ингибитором. Следовательно, у растений этого генотипа нормальная форма лепестков определяется не особыми генами (нормальной формы лепестков), а геном - подавителем гофрированности.
Обозначим ген гофрированности лепестков - А, нормальной формы - а (это основные гены формы лепестков), ингибитор гофрированности - I, ген отсутствия подавления - i. Тогда исходные формы с нормальными лепестками будут иметь генотипы IIАА и iiaа, гибриды F1 IiАа — также нормальные, а расщепление в F2 13/16 нормальных: 3/16 гофрированных можно представить как 9 (I-A-)+3 (I-аа) +1 (iiaa) = 13 нормальных и 3 iiA - гофрированных. Таким образом, подавление действия доминантного гена гофрированности лепестков доминантной аллелью другого гена (подавителя) обусловливает в F2 расщепление по фенотипу в отношении 13:3 [(9 + 3+1): 3].
Этот тип взаимодействия широко распространен в природе и наблюдается в наследовании окрашенности и неокрашенности зерен у кукурузы и оперения у кур и т. п. На рисунке 5 изображено наследование окраски луковицы у лука Allium сера.
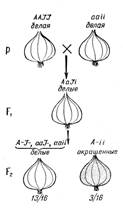
Рисунок 5. Наследование окраски луковицы у Allium сера (эпистаз): А - наличие окраски; а - отсутствие окраски; I - подавитель окраски; i - окраска не подавляется.
Расщепление 12:3:1. Доминантный эпистаз может давать и другое расщепление в F2 по фенотипу, а именно 12 : 3 : 1 [(9 + 3) : 3 : 1]. В этом случае, в отличие от предыдущего, форма, гомозиготная по обоим рецессивным генам, имеет специфический фенотип.
Например, некоторые собаки (Canis familiaris) с белой окраской шерсти при скрещивании с собаками, имеющими коричневую окраску, дают в F1 щенков с белой окраской, а в F2 расщепление на 12/16 белых, 3/16 черных и 1/16 коричневых (рис. 6). Если проанализировать это скрещивание отдельно по свойству окрашенности-неокрашенности и черно-коричневой окраске, то можно убедиться, что отсутствие окраски в F1 доминирует над ее наличием, а в F2 наблюдается расщепление 12:4, или 3:1. Расщепление на 3 черных и 1 коричневую свидетельствует о том, что черная окраска определяется доминантным геном, а коричневая - рецессивным. Теперь можно обозначить ингибитор окраски - I, его отсутствие - i, черную окраску - А, коричневую - а. Тогда легко представить генотипы исходных форм и гибридов. Подобный тип эпистаза встречается в наследовании окраски плодов у тыквы, окраски шерсти у овец (Ovis aries) и во многих других случаях. Таким образом, гены-подавители обычно не определяют сами какой-либо качественной реакции в развитии данного признака, а лишь подавляют действие других генов. Но в некоторых случаях это не так. Например, у хлопка (Gossypium) по окраске волокон в F2 наблюдается расщепление на 12 коричневых: 3 зеленых: 1 белую. Однако анализ коричневых волокон в ультрафиолетовых лучах позволяет выделить два типа коробочек: 3, имеющих волокна только с коричневым пигментом, и 9 — с коричневым и зеленым. У растений последнего типа зеленая окраска оптически не видна, так как коричневый пигмент ее как бы подавляет, т. е. является ингибитором.
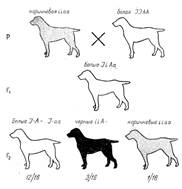
Рисунок 6. Наследование окраски шерсти у собак (эпистаз): А—черная окраска; а — коричневая; I — подавляет окраску; i — не подавляет.
Под рецессивным эпистазом понимают такой тип взаимодействия, когда рецессивная аллель одного гена, будучи в гомозиготном состоянии, не дает возможности проявиться доминантной или рецессивной аллели другого гена: аa>B- или aa>bb.
Расщепление 9:3:4 приводилось как пример комплементарного взаимодействия генов. Но эти же случаи можно рассматривать и как рецессивный эпистаз.
При скрещивании черных кроликов (AAbb) с белыми (ааВВ) все гибриды (АаВb) имеют окраску типа агути, а в F2 9/16 крольчат оказываются агути (А-В-), 3/16 черных (A-bb) и 4/16 белых (ааВ- и aabb). Эти результаты можно объяснить, предположив, что имеет место рецессивный эпистаз типа аа>В- и aa>bb. При этом кролики генотипа ааВ- и aabb оказываются белыми потому, что ген а в гомозиготном состоянии, блокируя образование пигмента, препятствует тем самым проявлению гена — распределителя пигмента В и гена черной окраски b.
Кроме описанных случаев одинарного рецессивного эпитаза, существуют и такие, когда рецессивная аллель каждого гена в гомозиготном состоянии одновременно реципрокно подавляет действие доминантной аллели комплементарного гена, т. е. аа эпистатирует над В-, bb над А-. Такое взаимодействие двух рецессивных подавителей называют двойным рецессивным эпистазом. В дигибридном скрещивании расщепление по фенотипу — 9 : 7, как и в случае комплементарного взаимодействия генов.
Следовательно, одно и то же расщепление можно трактовать как результат и комплементарного взаимодействия, и эпистаза. Один генетический анализ наследования при взаимодействии генов без знания биохимии и физиологии развития признака в онтогенезе не может раскрыть природы этого взаимодействия. Но без генетического анализа нельзя понять механизм наследственной детерминации развития этих признаков.
Полимерия. Рассмотренные до сих пор типы взаимодействия сенов относились к альтернативным, т. е. качественно различающимся, признакам.
Такие свойства организмов, как, например, темп роста и вес животного, длина стебля растения и т. п., нельзя разложить на четкие фенотипические классы; их необходимо измерять, взвешивать, подсчитывать, т.е. оценивать количественно. Подобные признаки обычно называют количественными или мерными признаками. Если расположить, например, овец одной породы в порядке возрастания их веса, то между самым мелким и крупным животными будет серия незаметных переходов, образующих непрерывный ряд.
Наследование таких признаков может происходить по-разному. При одном варианте, признак формируется под действием аллелей одного гена, который может быть представлен разным числом их в генотипе. Например, содержание витамина А в эндосперме зерна кукурузы зависит от количества доминантных аллелей гена у. Как известно, клетки эндосперма содержат три набора хромосом. Следовательно, путем скрещивания можно получить четыре различных по генотипу эндосперма кукурузы, содержащих разное количество доминантных и рецессивных аллелей у. Количество витамина А (в единицах активности) при разных дозах одного и того же гена оказывается следующим:
В эндосперме генотипа у у у . . . 0,05
» » » Y у у . . . 2,25
» » » Y Y у . . . 5,00
» » » Y У У . . . 7,50
Как видно из приведенных данных, действие одной дозы доминантного гена Y соответствует примерно 2,25 - 2,50 единиц активности витамина А. С увеличением дозы гена его действие суммируется, или кумулируется.
Такой тип действия гена называют кумулятивным или аддитивным, т. е. суммирующимся.
При другом варианте формирование количественного признака у организма, определяется взаимодействием многих доминантных генов, действующих на один и тот же признак или свойство. В этом случае количественные признаки, могут образовывать по своему проявлению непрерывный ряд. При этом, количественно варьирующий признак у разных особей одного и того же поколения будет определяться разным числом доминантных генов в генотипе. Так, при скрещивании рас пшениц (Triticum) с красными и белыми (неокрашенными) зернами шведский генетик Г. Нильсон-Эле в 1908 г. обнаружил в F2 обычное моногибридное расщепление в отношении 3:1.
Однако при скрещивании некоторых других линий пшениц, различающихся по таким же признакам, в F2 наблюдается расщепление в отношении 15/16 окрашенных : 1/16 белых. Окраска зерен из первой группы варьирует от темно- до бледно-красных (рис. 7).
Генетический анализ растений из семян F2 разных окрасок показал, что растения, выращенные из белых зерен и из зерен с наиболее темной (красной) окраской, в дальнейшем не дают расщепления. Из зерен с окраской промежуточного типа развились растения, давшие в последующих поколениях расщепление по окраске зерна. Анализ характера расщепления позволил установить, что в данном случае красную окраску зерен определяют доминантные аллели двух разных генов, а сочетание их рецессивных аллелей в гомозиготном состоянии определяет отсутствие окраски. Интенсивность окраски зерен зависит от числа доминантных генов в генотипе.

Рисунок 7. Наследование окраски зерна у пшеницы (полимерия)
Гены такого типа, одинаково влияющие на развитие одного признака, были названы генами с однозначным действием или полимерными. Такое же название получили и сами признаки. Поскольку эти гены однозначно влияют на один и тот же признак, было принято обозначать их одной латинской буквой с указанием номера разных генов: А1, А2, A3 и т. д. Этот тип взаимодействия генов получил название полимерии.
Следовательно, исходные родительские формы, давшие расщепление в F2: 15:1, имели генотипы А1А1А2А2 и а1а1а2а2. Гибрид F1: обладал генотипом Ala1A2a2, а в F2 появились зерна с разным числом доминантных генов. Наличие всех четырех доминантных аллелей A1A1A2A2 у 1/16 растений определяет самую интенсивную окраску зерна; 4/16 всех зерен имели три доминантных аллели (типа A1A1A2a2), 6/16 — две (типа A1a1A2a2), 4/16 — одну (типа A1a1a2a2), все эти генотипы определяли различные промежуточные окраски, переходные между интенсивно-красной и белой. Гомозиготной по обоим рецессивным генам (a1a1a2a2) являлась 1/16 всех зерен, и эти зерна оказались неокрашенными.
Нетрудно заметить, что частоты пяти перечисленных генотипических классов F2 распределяются в ряде: 1+4 + 6 + 4+1 = 16, который отображает изменчивость признака окраски зерна пшеницы в зависимости от числа доминантных аллелей в генотипе. Аналогичный тип наследования известен для некоторых видов окраски зерен кукурузы, колосковой чешуи у овса и т. п.
При накоплении доминантных полимерных генов их действие суммируется, т. е. они имеют кумулятивный эффект, поэтому взаимодействие такого типа называют кумулятивной полимерией.
Очевидно, что если у гибрида F1 число таких генов в гетерозиготном состоянии оказывается не два, а три А1а1А2a2А3а3 или более, то число комбинаций генотипов в F2 увеличивается. Этот ряд генотипов можно представить в виде биноминальной кривой изменчивости данного признака.
В опыте Нильсона-Эле тригибридное расщепление в F2 по генам окраски зерен пшеницы давало соотношение 63 красных к 1 неокрашенному. В F2: наблюдались все переходы от интенсивной окраски зерен с генотипом A1A1A2A2A3A3 до полного ее отсутствия у a1a1a2a2a3a3. При этом частоты генотипов с разным количеством доминантных генов распределялись в следующий ряд: 1+6+15 + 20+15 + 6+1=64. На рисунке 8 приведены гистограммы распределения частот генотипов с разным числом доминантных генов кумулятивного действия в моно-, ди-, три- и полигибридном скрещиваниях. Из этого сопоставления видно, что, чем большее число доминантных генов определяет данный признак, тем больше амплитуда изменчивости и тем более плавны переходы между различными группами особей.
Полимерно наследуется, например, пигментация кожи у человека. При бракосочетании негра и белой женщины рождаются дети с промежуточным цветом кожи (мулаты). У отца и матери мулатов могут родиться дети всех типов кожи с окраской разных оттенков, от черной до белой, что определяется комбинацией двух пар полимерных генов.

Рисунок 8. Распределение частот генотипов в F2 в случае кумулятивной полимерии.
Таким образом, при изучении наследования перечисленных выше признаков в F2 не наблюдается расщепления на определенные, легко отличимые фенотипические классы, как это имеет место в случае альтернативных признаков: гладкая или морщинистая форма семян у гороха и т. д. Полимерные признаки, как правило, необходимо измерять или подсчитывать. Поэтому, в отличие от альтернативно наследующихся, так называемых качественных признаков, их называют количественными признаками. При наследовании таких признаков потомство гибрида по фенотипическому проявлению образует непрерывный ряд.
В принципе деление признаков на количественные и качественные условно. Как те, так и другие признаки можно и должно измерять при изучении их наследования, поскольку без количественной оценки любого явления природы не может быть объективного его анализа.
В качестве примера приведем результаты скрещивания двух форм кукурузы — длиннопочатковой и короткопочатковой. Как видно из рисунка 9, початки по их длине у исходных линий кукурузы № 60 (короткопочатковая) и № 54 (длиннопочатковая), а также у гибридов первого и второго поколений распределяются с определенной закономерностью. Нетрудно заметить, что эти две линии сильно различаются между собой, но в пределах каждой из них длина початков колеблется незначительно. Это указывает на то, что они наследственно сравнительно однородны. Захождения в размерах початков родительских форм нет. У гибридных растений F1 длина початков оказывается промежуточной, с небольшой изменчивостью. В F2 размах изменчивости значительно увеличивается. Следовательно, непрерывный ряд изменений по длине початка кукурузы можно представить как ряд генотипов с различным числом доминантных генов, обусловливающих данный количественный признак.
Тот факт, что при небольшом числе исследованных растений второго поколения у некоторых из них воспроизводится длина початков, свойственная родительским формам, может указывать на участие небольшого числа полимерных генов в определении длины початка у скрещиваемых форм. Такое предположение вытекает из известной нам формулы 4n , определяющей число возможных комбинаций гамет, образующих зиготы в F2, в зависимости от числа пар генов, по которым различались исходные родительские формы. Появление в опыте среди 221 растения F2 форм, сходных с родительскими, указывает на то, что число независимо наследующихся генов, определяющих длину початка, не должно превышать трех (43 = 64) или четырех (44 = 256). Большая изменчивость признака указывает на его сложную генетическую обусловленность, а меньшая — на меньшее число факторов, его определяющих.
Приведенные примеры анализа наследования количественных признаков иллюстрируют лишь один из возможных путей их изучения. Другой путь — применение математических методов. Анализ наследования количественных признаков и действия полимерных генов чрезвычайно сложен.

Рисунок 9. Наследование длины початков (в см) у кукурузы (полимерия).
Изучение полимерных генов имеет не только теоретический, но и большой практический интерес. Хозяйственно ценные признаки у животных и растений, такие, как жирномолочность коров, яйценоскость кур, длина колоса пшеницы, содержание сахара в корнеплодах свеклы и многие другие, наследуются по типу полимерии.
Проявление полимерных признаков в очень большой степени определяется условиями развития организма. Так, молочная продуктивность коров, длина шерсти овец, скорость роста свиней во многом зависят от условий кормления и содержания животных. Величина клубней картофеля, початков кукурузы или длина стебля льна определяются в значительной мере качеством вносимых удобрений, количеством осадков и т. п.
Некумулятивная полимерия. Полимерные гены с однозначным действием могут определять и качественные, т. е. альтернативные, признаки. Примером может служить наследование оперенности ног у кур (Gallus gallus). От скрещивания пород, имеющих оперенные и неоперенные ноги, в F1 появляются цыплята с оперенными ногами. Во втором поколении происходит расщепление по фенотипу,в отношении 15/16 с оперенными ногами и 1/16 с неоперенными, т. е. наблюдается два фенотипичееких класса (рис. 10),
Очевидно, порода с оперенными ногами гомозиготна по двум парам однозначных доминантных генов (A1A1A2A2), а с неоперенными — имеет генотип а1а1а2а2. Сочетание гамет при оплодотворении дает гибриды с генотипом А1а1А2а2. Доминантные аллели каждого из двух генов действуют качественно однозначно, т. е. определяют оперенность ног. Поэтому генотипы А1-А2- (9/16), A1-a2a2 (3/16) и a1a1A2-(3/16) соответствуют фенотипу с оперенными ногами, а генотип а1а1а2а2 (1/16) с неоперенными.
Таким же образом осуществляется наследование формы стручка у пастушьей сумки (Capsella bursa pastoris). При скрещивании расы, имеющей яйцевидные стручки, с расой, у которой плоды треугольной формы, в F1 все растения имеют треугольные стручки, а в F2 наблюдается расщепление в отношении 15: 1 [(9+3 + 3) : 1].
В двух приведенных примерах наличие в генотипе разного количества доминантных полимерных генов однозначного действия не изменяет выраженности признака. Достаточно одной доминантной аллели любого из двух генов, чтобы вызвать развитие признака. Поэтому такой тип взаимодействия генов был назван некумулятивной полимерией.

Рисунок 10. Наследование оперенности ног у кур (полимерия): А - оперенные ноги, а - неоперенные.
Все рассмотренные типы взаимодействия генов: комплементарное, эпистатическое и полимерное видоизменяют классическую формулу расщепления по фенотипу (9: 3: 3 : 1), установленную Менделем для дигибридного скрещивания. В таблице 6 приведены некоторые типы расщепления по фенотипу для дигибридного скрещивания, при этом все они показаны с точки зрения доминантного и рецессивного эпистаза.
Таблица 6
Соотношение фенотипичееких классов расщепления в потомстве дигетерозиготы при некоторых типах взаимодействия генов
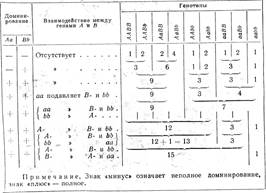
Все приведенные типы расщепления по фенотипу столь же закономерны, как 9:3:3: 1; они являются не следствием нарушения генетического механизма расщепления, а результатом взаимодействия генов между собой в индивидуальном развитии.
Модифицирующее действие генов. При изучении явления взаимодействия были открыты гены основного действия, т. е. такие, которые определяют развитие признака или свойства, например выработку пигментов, форму цветка и т. п., и такие, которые сами по себе не определяют какую-либо качественную реакцию или признак, а лишь усиливают или ослабляют проявление действия основного гена. Это гены-модификаторы, а их действие — модифицирующее.
Одни из генов-модификаторов могут усиливать эффект основного гена, другие ослаблять. Например, у крупного рогатого скота пегая окраска шерстного покрова определяется рецессивным геном и двумя модификаторами, ослабляющими или усиливающими эффект основного гена пегости (рис. 11). Независимо от наличия или отсутствия модификаторов, при скрещивании животного, имеющего сплошную окраску, с пегим в F1 будет доминировать сплошная окраска, а в F2 — осуществляться расщепление 3:1. Действие модификаторов обнаруживается в присутствии гена пегости и проявляется в увеличении или уменьшении непигментированных участков шерстного покрова.



Рисунок 11. Модификация пегости у крупного рогатого скота: 1—усиленная пегость; 2 — пегость; 3 — ослабленная пегость.
Дата добавления: 2016-04-02; просмотров: 5349;