ВЫСОКОСПЕЦИФИЧНЫЕ ПРОТЕИНАЗЫ
Адсорбционный центр этих ферментов имеет сложное строение. Они способны распознавать структуру радикала не только одной из аминокислот, но целого участка полипептидной цепи, в составе которого находится пептидная связь, которую гидролизует фермент. Часто высокоспецифичная протеиназа может узнать и гидролизовать только одну связь из сотен других, имеющихся в белке-субстрате. Такое высокоспецифичное расщепление молекулы белка в одном строго определенном месте называется “ограниченный протеолиз”.
Высокоспецифичные протеиназы можно разделить на две группы:
1. Внутриклеточные высокоспецифичные протеиназы. Обеспечивают постсинтетическую модификацию белка. Молекулы белка синтезируется в рибосомах в виде единственного полипептида, в составе которого гораздо больше аминокислот, чем в том белке, который затем из него образуется.
Постсинтетическая модификация белка включает в себя множество разнообразных процессов, которые различны для каждого отдельного белка.
Например, может происходить химическая модификация некоторых аминокислотных радикалов (так, пролин в составе коллагена превращается в оксипролин).
После синтеза белка к нему присоединяются углеводные фрагменты. Так образуются гликозилированнные белки. Постсинтетические превращения, которые сопровождают ограниченный протеолиз, называются "ПРОЦЕССИНГ БЕЛКА".
Все реакции процессинга можно разделить на две фазы:
а) отщепление "сигнального" пептида;
б) последующая постсинтетическая модификация.
Обычно белки синтезируются так, что на N-конце такого белка имеется последовательность от 15 до 30 аминокислотных фрагментов, которые не входят в состав окончательного белка. Оказалось, что в такой СИГНАЛЬНОЙ ПОСЛЕДОВАТЕЛЬНОСТИ (или сигнальном пептиде очень много аминокислот с гидрофобными радикалами. Поэтому сигнальная последовательность очень устойчива к действию протеолитических ферментов. Гидрофобность сигнальной последовательности обеспечивает молекуле белка проникновение через мембраны.
Выделяют три главные функции сигнальных пептидов:
а) обеспечивают устойчивость синтезированного белка к протеолизу на всем пути этого белка от рибосом до места, где белок выполняет свою функцию в клетке;
б) создают условия для переноса белка через мембраны.
Таким образом, сигнальные пептиды обеспечивают транспорт белка от места синтеза к месту назначения - обеспечивают адресную функцию.
Даже после отщепления сигнального пептида формирование окончательного белка еще на закончено: остается длинная полипептидная цепь, которая еще должна быть укорочена.
Опять протекает серия реакций ограниченного протеолиза, в результате которых полипептидная цепь укорачивается по-разному: иногда происходит укорочение путем гидролиза со стороны С-конца; иногда гидролиз происходит со стороны N-конца; в некоторых случаях расщепление полипептида происходит в середине цепи в результате гидролиза в двух местах.
ПРИМЕРЫ РАБОТЫ ВНУТРИКЛЕТОЧНЫХ ВЫСОКОСПЕЦИФИЧНЫХ ПРОТЕИНАЗ
Пример 1: СОЗРЕВАНИЕ МОЛЕКУЛЫ ГОРМОНА ИНСУЛИНА:

|
Созревшая молекула состоит из двух полипептидных цепей, которые соединены двумя дисульфидными связями. В одной цепи (А-цепь) содержится 21 аминокислотный остаток, а во второй (В-цепь) - 30 аминокислотных остатков.
Оказалось, что этот белок синтезируется в виде единственной полипептидной цепи (ПРЕПРОИНСУЛИН), в которой содержится 100 аминокислотных остатков. После гидролиза со стороны N-конца от молекулы отрывается сигнальный пептид (16 аминокислот) и образуется ПРОИНСУЛИН. Наличие сигнальной последовательности в препроинсулине позволяет ему проникать через мембраны трубочек эндоплазматического ретикулума. А превращение препроинсулина в проинсулин происходит внутри трубочек под действием высокоспецифичной протеиназы.
Затем, в аппарате Гольджи начинается и в секреторных гранулах завершается вторая группа реакций процессинга. В ходе этих реакций образуется В-цепь, а затем со стороны С-конца на расстоянии в 20 аминокислотных фрагментов от конца происходит гидролиз связи между арг79 и гли80. В конечном счете от молекулы проинсулина отделяется 33-членный срединный пептид. В результате образуется ИНСУЛИН.
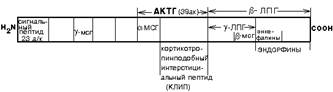
Пример 2. СОЗРЕВАНИЕ МОЛЕКУЛЫ АДРЕНОКОРТИКОТРОПНОГО ГОРМОНА ГИПОФИЗА (АКТГ).
Белок кортикотропин синтезируется в составе более крупной молекулы, которая содержит 264 аминокислотных фрагмента и называется ПРООПИОКОРТИН.
|
Сам АКТГ состоит из аминокислот от 131-й до 170-й в составе этого белка, а остальные участки содержат сигнальную последовательность и в составе этой же молекулы содержится полипептид, из которого образуется меланоцитстимулирующий гормон (МСГ). В ходе процессинга от проопиокортина вначале отщепляется сигнальная последовательность и затем после двух реакций протеолиза со стороны N-конца и С-конца отделяется пептид гамма2-МСГ (меланоцитстимулирующий гормон). АКТГ освобождается с N-конца.
КЛИП - кортикотропинподобный пептид промежуточной доли гипофиза. На С-концевой части этого пептида содержатся последовательности, из которых образуются ЭНДОРФИНЫ (эндогенные морфины). По структуре они являются пептидами. Например, пептид скотофобин вызывает у животных боязнь темноты (даже если животным, ведущим ночной образ жизни, его ввести, то они начинают бояться темноты).
Внутриклеточные протеиназы, которые обеспечивают реакции процессинга, обладают высокой субстратной специфичностью. Каждая такая протеиназа действует на один определенный белок, а следующая протеиназа действует только на продукт первой реакции.
Совсем по другому организованы системы внеклеточных протеиназ.
ВНЕКЛЕТОЧНЫЕ ПРОТЕИНАЗЫ
Примером может служить система свертывания крови. Это совокупность более чем десяти разных белков. Многие из этих белков являются неактивными формами протеиназ (т.е. проферментами). Например, среди белков системы свертывания крови имеется фактор XII. Он вступает в контакт с чужеродной поверхностью (например, со стеклом при повреждении стенки сосуда) и при этом происходят конформационные изменения. На поверхность молекулы выступает ее активный центр, который был раньше спрятан. Этот белок уже становится активным и может разрушать одну пептидную связь в другом белке, который тоже в результате этого воздействия из профермента превращается в активный фермент.
Для этого активного фермента субстратом является следующий белок плазмы, который превращается под действием второго звена из профермента в активный фермент, пока процесс не дойдет до фибриногена. Очередной протеолитический фермент действует на фибриноген, превращая его в фибрин. Фибрин - не является ферментом, он плохо растворим в воде, образует нити, которые выпадают в осадок. В этом осадке запутываются форменные элементы крови. Так образуется кровяной сгусток.
Именно высокая субстратная специфичность позволяет протеиназам плазмы образовать в крови систему, звенья которой работают строго последовательно. Эта система - система свертывания крови работает по принципу каскадности. Происходит постепенное усиление первоначально слабого сигнала. Свертывание крови происходит постоянно, но оно уравновешивается процессом фибринолиза. Это обеспечивается наличием в плазме крови фермента плазмина, который образуется из плазминогена и не является звеном каскада свертывания. Плазмина, который содержится в крови, достаточно, чтобы обеспечить гидролиз фибрина внутри сосудов. При нарушениях фибринолиза наблюдается ДВС-синдром (синдром диссеминированного внутрисосудистого свертывания).
К протеолитическим системам плазмы крови относятся также СИСТЕМА КОМПЛЕМЕНТА и СИСТЕМА РЕГУЛЯЦИИ СОСУДИСТОГО ТОНУСА (с помощью вазоактивных пептидов). Подробно об этих системах, а также о работе системы свертывания крови изложено в лекции “ПРОТЕОЛИТИЧЕСКИЕ СИСТЕМЫ КРОВИ”.
Дата добавления: 2016-03-10; просмотров: 798;