Рестриктазы, сайты рестрикции
Гексануклеотидные последовательности
 |
|
|
|
 |
|
|
|
 EcoR1 GAATTC 5’-G AATTC-3’
EcoR1 GAATTC 5’-G AATTC-3’
5’-выступающие концы, «липкие»
 |
|
|
|
3’-выступающие концы, «липкие»
 |
Csp6I GTAC тетрануклеотидные последовательности
CATG
 |
AsuII TTCGAA
 AAGCTT
AAGCTT
Направленный мутагенез с помощью олигонуклеотидов
5’- GGC ATC GGG TAC TAC ATC –3’ ДНК

 внесение
внесение
делеции GGC ATC TAC TAC ATC Олиг

 5’- GGC ATC GGG TAC TAC ATC –3’ ДНК
5’- GGC ATC GGG TAC TAC ATC –3’ ДНК
 внесение
внесение
вставки ATC GGG CCC TAC TAC Олиг
5’- GGC ATC GGG TAC TAC ATC –3’ ДНК
Gly Ile Gly Tyr Tyr Ile
Замена
аминокислоты ATC GGG АAC TAC Олиг
Ile Gly Asn Tyr
5’- GGC ATC GGG TAC TAC ATC –3’ ДНК
 |
внесение ATC GGG TAC СAC Олиг
 уникального
уникального
сайта Kpn1
рестрикции
После смыкания интересующего фрагмента ДНК (гена) с генетическим вектором с помощью ДНК-лигазы образованные рекомбинантные молекулы вводят в клетки с целью добиться их репликации (за счет генетического вектора) и увеличения количества копий. Наиболее популярным способом введения в клетки рекомбинантных молекул ДНК, в которых вектором служит плазмида, является трансформация Е. coli. С этой целью бактериальные клетки предварительно обрабатывают кальцием или рубидием (ионами) для того, чтобы они стали «компетентными» в восприятии рекомбинантной ДНК.
Чтобы повысить частоту проникновения ДНК в клетки, используют метод электропорации, заключающийся в кратком экспонировании клеток в интенсивном электрическом поле. Эта обработка создает полости в мембранах клеток, что способствует лучшему восприятию клетками ДНК- После введения рекомбинантных молекул ДНК в бактерии последние высевают на МПА, обогащенный антибиотиками для селекции желаемых клеток, т. е. клеток, содержащих рекомбинантные молекулы ДНК. Частота трансформации является невысокой. Обычно один трансформант возникает на 106 высеянных клеток. Если же вектор является фаговым, то прибегают к трансфекции клеток (бактерий или дрожжей) фагом. Что касается соматических клеток животных, то их трансфекцию осуществляют ДНК в присутствии химических веществ, облегчающих прохождение ДНК через плазматические мембраны. Возможны также прямые микроинъекции ДНК в овоциты лягушек, в культивируемые соматические клетки и в эмбрионы млекопитающих.
Важнейшим моментом, связанным с молекулярным клонированием, является поиск способа, позволяющего установить, действительно ли клонируемый фрагмент включился в вектор и вместе с вектором, образовав рекомбинантную молекулу ДНК, вошел в клетки. Если речь идет о бактериальных клетках, то один из способов основан на учете инсерционной инактивации плазмидного (векторного) гена резистентности. Например, в плазмидном векторе pBR322, детерминирующем резистентность к ампициллину и тетрациклину, единственный сайт для рестриктазы Pst I находится в локусе, занимаемом геном резистентности к ампициллину. Pst I —плавление на этом сайте генерирует липкие концы, позволяющие лигирование клонируемого фрагмента с векторной ДНК. Однако при этом плазмидный (векторный) ген ампициллинрезис-тентности инактивируется, тогда как ген тетрациклинрезистентнос-ти на векторе остается интактным. Именно, ген тетрациклинрезистентности и используется для селекции клеток, трансформируемых рекомбинантными молекулами ДНК. Чтобы убедиться, что клетки выросших колоний на среде с тетрациклином действительно содержат рекомбинантные молекулы ДНК, их проверяют с помощью так называемого «спот-теста» на паре чашек с плотной средой, одна их которых содержит ам-пициллин, тогда как другая лишена этого антибиотика. Клонируемые ДНК содержатся лишь в трансформантах, резистентных к тетрациклину Что касается трансформантов, резистентных одновременно к ампициллину и тетрациклину (АрТс), то они содержат плазмидные (векторные) молекулы, которые спонтанно приобрели кольцевую форму без включения в них чужеродной (клонируемой) ДНК. Другой способ обнаружения инсерции чужеродных (клонируемых) фрагментов в плазмидный вектор основан на использовании вектора, содержащего ген галактозидазы. Инсерция чужеродной ДНК в этот ген неизбежно инактивирует синтез р-галактозидаза, что может быть обнаружено посевом трансформированных клеток на среду, которая содержит субстраты галактозидазы. Эта среда позволяет вести селекцию окрашенных колоний клеток.
Совокупность колоний-трансформантов, выросших на селективной среде, представляет собой совокупность клеток, содержащих клоны разных фрагментов (генов) клонируемой геномной или кДНК. Коллекции этих клонов формируют так называемые библиотеки ДНК, широко используемые в генно-инженерных работах.
Заключительной стадией клонирования генов является выделение и исследование клонированной ДНК, включая секвенирование. Перспективные штаммы бактерий или соматических клеток, содержащих рекомбинантные молекулы ДНК, которые контролируют синтез интересующих белков, имеющих коммерческую ценность, передают в промышленность.
Методы конструирования гибридных молекул ДНК in vitro. Способы введения ДНК в клетку (трансформация, трансфекция, электропорация)
Способы прямого введения генов в клетку
Прямое введение гена в клетку осуществляют несколькими способами:
1. Трансфекция
2. Микроинъекция
3. Электропорация
4. Метод «мини-клеток»
5. Упаковка в липосомы
6. Электронная пушка
При трансфекции ДНК адсорбируется на кристаллах фосфата кальция (Грэхем Ван дер Эб, 1973). Образуются частицы кальциевого преципитата. Они поглощаются клеткой путем фагоцитоза.
Для повышения эффективности трансформации к специфической ДНК, содержащей ген, по которому будет производиться селекция, добавляется неспецифическая ДНК-носитель. Обычно для этой цели берут ДНК из тимуса теленка или спермы лосося. Часть ДНК связывается с мембраной и не попадает в клетки. ДНК акцептируют от 15 до 90% клеток. Через несколько суток после введения небольшая доля клеток способны экспрессировать чужеродные гены, но затем уровень экспрессии падает и более или менее стабильную трансформацию претерпевает 10-3 - 10-5 клеток.
Для трансфекции используется и ДЭАЭ-декстран, полимер, адсорбирующий ДНК. Эффект вхождения в клетки и время экспрессии высоки, но частота стабильной трансформации ниже, чем при использовании преципитата кальция. Частоту трансфекции увеличивает глицериновый шок (4 минуты в 15% растворе глицерина в НEPES-буфере).
В клетки можно вводить любой ген, если заранее лигировать его с клонированным селективным маркером. Однако дальнейшие исследования показали, что лигирование вне клетки не обязательно. Клетки, поглощающие селективный ген, вместе с ним поглощают и другую ДНК, имеющуюся в кальциевом преципитате. Таким образом, пользуясь методом котрансформации, практически любой клонированный сегмент ДНК можно ввести в культивируемые клетки эукариот, если включить эту ДНК вместе с селективным маркером в состав смеси для образования кальциевого преципитата.
Для трансфекции можно использовать хромосомы или фрагменты хромосом. Клетки-доноры блокируются на стадии митоза. Митотические хромосомы высвобождаются под воздействием осмотического шока и гомогенизации. Их очищают путем дифференциального центрифугирования. Хромосомы осаждают на поверхности клеток хлористым кальцием, а через несколько часов обрабатывают реагентом, способным перфорировать мембраны (например, глицерином).
Для обработки клеток-рецепиентов используются грубо очищенные препараты хромосом, так как хромосомы при этом разрушаются меньше всего. Количество хромосом для обработки 1 клетки ограничено. Лучше использовать не более 20 хромосом на 1 клетку-рецепиент, так как при высоких концентрациях хромосом в суспензии они агглютинируют. Рецепиентная клетка содержит фрагменты донорных хромосом, которые могут встраиваться в геном, могут реплицироваться самостоятельно. Во введенных фрагментах часто наблюдаются делеции.
Не все клетки способны к трансформации геномной ДНК с высокой частотой. Человеческие фибробласты эффективно включают плазмидную ДНК и почти не включают геномную.
Микроинъекция ДНК в клетки млекопитающих стала возможной с появлением прибора для изготовления микропипеток диаметром 0.1-0.5 микрона и микроманипулятора (рис. 9.1). Так, плазмиды, содержащие фрагмент вируса герпеса с геном тимидинкиназы (ТК) и плазмиду рВR322, были инъецированы в ТК--клетки и было показано, что ТК-ген проник в ядра и нормально в них реплицировался. Метод введения ДНК с помощью микроинъекций был разработан в начале 70-х годов Андерсоном и Диакумакосом. В принципе, при наличии хорошего оборудования можно за 1 час инъецировать 500-1000 клеток, причем в лучших экспериментах в 50% клеток наблюдается стабильная интеграция и экспрессия инъецированных генов. Преимущество описываемого метода заключается также в том, что он позволяет вводить любую ДНК в любые клетки, и для сохранения в клетках введенного гена не требуется никакого селективного давления.
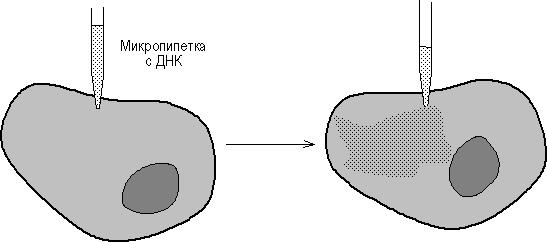
Рис. 9.1 - Введение ДНК путем микроинъекции
Электропорация основана на том, что импульсы высокого напряжения обратимо увеличивают проницаемость биомембран. В среду для электропорации добавляют клетки и фрагменты ДНК, которые необходимо ввести в клетки (рис. 9.2). Через среду пропускают высоковольтные импульсы (напряжение 200 - 350 В, длительность импульса 54 мс), приводящие к образованию пор (электропробой) в цитоплазматической мембране, время существования и размер которых достаточны, чтобы такие макромолекулы, как ДНК, могли из внешней среды войти в клетку в результате действия осмотических сил. При этом объем клетки увеличивается.
Напряженность электрического поля и продолжительность его действия, концентрации трансформирующей ДНК и реципиентных клеток для каждой системы клеток подбирают экспериментально, с тем чтобы достичь высокого процента поглощения ДНК выжившими клетками. Показано, что в оптимальных условиях электропорации количество трансформантов может достигать 80% выживших клеток.
Электропорация — физический, а не биохимический метод, и это, по-видимому, обусловливает его широкое применение. Многочисленные исследования продемонстрировали, что электропорация может успешно использоваться для введения молекул ДНК в разные типы клеток, такие как культивируемые клетки животных, простейшие, дрожжи, бактерии и протопласты растений. Электропорирующий эффект высоковольтного разряда на бислойную липидную мембрану, по-видимому, зависит от радиуса ее кривизны. Поэтому мелкие бактериальные клетки эффективно поглощают ДНК при значительно большей напряженности (10 кВ/см и более), чем крупные животные и растительные клетки, эффективно поглощающие ДНК при напряженности поля 1—2 кВ/см.
Электропорация — наиболее простой, эффективный и воспроизводимый метод введения молекул ДНК в клетки. Однако до недавнего времени этот метод использовался в ограниченном числе лабораторий в связи с отсутствием серийных приборов — электропораторов. Появление и совершенствование таких приборов в ближайшие годы приведет к широкому применению данного подхода в генетической инженерии самых разных типов клеток.
 Рис. 9.2 - Метод электропорации
Рис. 9.2 - Метод электропорации
«Мини-клетки» получают путем блокирования донорных клеток митозе колцемидом. При продолжительной обработке клеток колцемидом в них вокруг каждой хромосомы формируется новая ядерная мембрана. Обработка цитохалазином В и центрифугирование приводит к образованию мини-клеток, представляющих микроядра, инкапсулированные в цитоплазматическую мембрану.
Полученные мини-клетки очень чувствительны к разного рода воздействиям, поэтому для слияния подбирают специальные мягкие условия. Метод трудный, капризный, эффективность низкая – 10-6 – 10-7.
Упаковка в липосомы используется для защиты экзогенного генетического материала от разрушающего действия рестриктаз.
Липосомы - сферические оболочки, состоящие из фосфолипидов. Получают их путем резкого встряхивания смеси водного раствора и липидов, либо обрабатывая ультразвуком водные эмульсии фосфолипидов. Липосомы, состоящие из фосфатидилсерина и холестерина наиболее пригодны для введения ДНК в клетки животных и растений. Системы переноса с помощью липосом низкотоксичны по отношению к клеткам.
Метод биологической баллистики (биолистики) является одним из самых эффективных на сегодняшний день методов трансформации растений, особенно однодольных.
Суть метода заключается в том, что на мельчайшие частички вольфрама, диаметром 0,6—1,2 мкм, напыляется ДНК вектора, содержащего необходимую для трансформирования генную конструкцию. Вольфрамовые частички, несущие ДНК, наносятся на целлофановую подложку и помещаются внутрь биолистической пушки. Каллус или суспензия клеток наносится в чашку Петри с агаризированной средой и помещается под биолистическую пушку на расстоянии 10—15 см. В пушке вакуумным насосом уменьшается давление до 0,1 атм. В момент сбрасывания давления вольфрамовые частички с огромной скоростью выбрасываются из биолистической пушки и, разрывая клеточные стенки, входят в цитоплазму и ядро клеток.
Обычно клетки, располагающиеся непосредственно по центру, погибают из-за огромного количества и давления вольфрамовых частиц, в то время как в зоне 0,6—1 см от центра находятся наиболее удачно протрансформированные клетки. Далее клетки осторожно переносят на среду для дальнейшего культивирования и регенерации.
С помощью биолистической пушки были протрансформированы однодольные растения, такие, как кукуруза, рис, пшеница, ячмень. При этом были получены стабильные растения-трансформанты. Кроме успехов в получении трансгенных однодольных, биолистическая трансформация применяется для прямого переноса ДНК в эмбриогенную пыльцу и дальнейшего быстрого получения трансгенных дигаплоидных растений, которые являются важным этапом в селекционной работе. В настоящее время этим методом была проведена трансформация растений табака и после регенерации гаплоидных растений получены стабильные трансформанты.
Трансформация.
В настоящее время бактерия E. coli является самой изученной клеткой из всех существующих. У большинства наиболее полно изученных фагов клеткой - хозяином является также E. coli.
Протопласт E. coli одет в муреиновый мешок, прилегающий к внешней мембране. E. coli относится к микроорганизмам, не обладающим физиологической компетентностью к поглощению экзогенной ДНК. Поэтому необходимо создать условия, позволяющие преодолеть барьер клеточной стенки. Сначала получают сферопласты путем обработки клеток лизоцимом в изотоническом растворе.
Липополисахаридный слой внешней мембраны грамотрицательной бактерии стабилизирован двухвалентными катионами, поэтому для разрыхления внешней мембраны E. coli используется комплексообразователь этилендиаминтетрауксусная кислота (EDTA), которая связывает двухвалентные катионы. При обработке EDTA часть липополисахаридов высвобождается из внешней мембраны клетки, и лизоцим может достигнуть муреинового мешка и гидролизовать его. Это ведет к повышению проницаемости клеточной оболочки. Усовершенствование методов получения сферопластов E. coli и их трансфекции позволили достичь достаточно высокой эффективности трансформации молекулами ДНК различных фагов.
Обнаружено, что на инфекционность существенное влияние оказывает форма молекул фаговых ДНК, которую они принимают in vivo. Фаги с кольцевой или линейной, но быстро замыкающейся ДНК (лямбоидные фаги) характеризуются наибольшей эффективностью трансфекции.
Успешное проведение экспериментов на кишечной палочке стало стимулом для проведения аналогичных исследований с другими прокариотическими организмами. Наибольших успехов удалось достичь с клетками Bacillus subtilis. B. subtilis - непатогенный почвенным микроорганизм, растущий в строго аэробных условиях. Бациллы не образуют токсинов и непатогенны ни для животных, ни для человека, тогда как клеточная стенка E. coli содержит эндотоксин, который довольно трудно отделить от продуктов генной инженерии. Кроме того, клеточная стенка бацилл имеет простую структуру и бактерии могут секретировать многие белки в культуральную жидкость. 20 различных видов бацил секретируют в культуральную жидкость более 40 ферментов с внеклеточной локализацией. E. coli секретирует в среду относительно мало белков, а выделение и очистка их затруднены. В бациллах также обнаружены плазмиды и фаги, которые к настоящему моменту уже хорошо изучены.
Чужеродные гены клонируют в так называемых челночных векторах. Эти вектора с одинаковым успехом реплицируются в клетках нескольких хозяев, в данном случае, в клетках E. coli и B.subtilis. Векторы были получены комбинацией in vitro фрагментов этих плазмид.
Гены E. coli со своими регуляторными районами не функционируют в B.subtilis, поэтому были использованы собственные гомологичные районы B.subtilis.
Для конструирования рекомбинантной ДНК, содержащей в своем составе ген, который должен экспрессироваться, придерживаются следующей стратегии. Синтезируют кДНК или из клонотеки выделяют клетки, несущие фрагмент генома с нужным геном, и клонируют их в соответствующем векторе. Фрагменты геномной ДНК подвергают модификации - удаляют из них некодирующие области и участки соседних генов. Часто для проведения этой операции необходимо секвенирование данного фрагмента ДНК. Затем конструируются промежуточные рекомбинантные ДНК, в которых ген помещается под контроль бактериальных регуляторных элементов (промотор, оператор, точка связывания с рибосомами). Эти регуляторные элементы выделяют из гибридных плазмид, сконструированных специально как источники регуляторных элементов. Полученная конструкция встраивается в подходящий вектор, например, pBR 322, и ген экспрессируется в бактериальной клетке.
Однако удобнее встраивать ген в специальный вектор для экспрессии, который уже содержит регуляторные элементы, обеспечивающие активную экспрессию после введения рекомбинантной плазмиды в бактериальную клетку. К таким эффективным регуляторным участкам относится, например, сильный промотор гена бэта-лактамазы (ген устойчивости к пенициллину, входящий в состав плазмиды pBR 322). Ряд генов, в том числе и ген инсулина, встраивали в сайт рестрикции Pst I, который расположен в структурной части гена. Промотор этого гена обеспечивает эффективную транскрипцию, которая продолжается до тех пор, пока РНК-полимераза не дойдет до сигнала терминации встроенного гена.
В качестве примера маркирования вектора могут служит первые эксперименты с E. coli, а точнее с одной из ее плазмид рBR322, проведенные Гилбертом для получения инсулина. Плазмида pBR322 содержит 2 гена, которые определяют устойчивость к ампициллину и тетрациклину. Рестриктаза PstI расщепляет плазмиду в средней части гена, кодирующего фермент устойчивости к апициллину. После расщепления плазмиды на ее концы с помощью концевой трансферазы надстраивали последовательность из четырех нуклеотидов с остатками гуанина. Затем, как обычно, с помощью лигаз "вшивали" ген проинсулина, получая рекомбинантную ДНК. Встроенный в плазмиду фрагмент ДНК нарушал синтез фермента, разрушающего ампициллин, но ген, обеспечивающий устойчивость к тетрациклину, оставался активным. Трансформированные таким образом клетки E. coli синтезировали гибридный белок, содержащий последовательности пенициллазы и проинсулина, поэтому биологически активный инсулин получали путем отщепления пенициллазы и средний сегмент проинсулина.
С другой стороны, если фрагмент чужеродной ДНК встраивается в один из генов устойчивости, то последний инактивируется. Следовательно, успешное встраивание фрагмента чужеродной ДНК в один из этих генов легко детектировать по исчезновению у бактерий устойчивости к данному антибиотику.
Характеристика векторов для переноса генов в животные клетки
Манипуляции с клетками млекопитающих можно разделить на 2 большие группы: эксперименты с соматическим клетками и эксперименты по трансформации половых клеток. В последнем случае конечный результат – получение трансгенных организмов.
Характеристика векторов для переноса генов в животные клетки.
Одними из лучших носителей для введения чужеродной информации в животную клетку являются вектора на основе ретровирусов, например, на основе вируса лейкоза мышей. Они обеспечивают высокоэффективный перенос генов и их стабильное встраивание в хромосому клеток-мишеней. В основном трансформации животных клеток осуществляют либо с помощью ретровирусов (около 40% от всех трансформаций), либо путем упаковки ДНК в липосомы (25%), реже используют аденовирусы, так как они могут вызывать сильный иммунный ответ, кроме того, невозможно их повторное введение.
Если проблема доставки чужеродной ДНК in vitro практически решена, а ее доставка в клетки-мишени разных тканей in vivo успешно решается (главным образом путем создания конструкций, несущих рецепторные белки, в том числе и антигены, специфичные для тех или иных тканей), то другие характеристики существующих векторных систем - стабильность интеграции, регулируемая экспрессия, безопасность — все еще нуждаются в серьезных доработках.
Прежде всего это касается стабильности интеграции. До настоящего времени интеграция в геном достигалась только при использовании ретровирусных либо аденоассоциированных векторов. Повысить эффективность стабильной интеграции можно путем совершенствования генных конструкций типа рецептор-опосредованных систем, либо путем создания достаточно стабильных эписомных векторов (то есть ДНК-структур, способных к длительной персистенции внутри ядер).
В последнее время особое внимание уделяется созданию векторов на базе искусственных хромосом млекопитающих (MAC - mammalian artificial chromosomes). Благодаря наличию основных структурных элементов обычных хромосом такие мини-хромосомы длительно удерживаются в клетках и способны нести полноразмерные (геномные) гены и их естественные регуляторные элементы, которые необходимы для правильной работы гена, в нужной ткани и в должное время. Такие искусственные хромосомы уже созданы для дрожжей (YAK), так как геном дрожжей полностью картирован.
Для идентификации модифицированных клеток, необходимы маркеры. Если трансформируют соматические клетки, то применяют обычно селективные маркеры. Аксель с коллегами из колледжа терапии и хирургии Колумбийского университета исправили таким образом генетический дефект клеток мыши. Они взяли фрагмент ДНК, содержащий ген тимидинкиназы (ТК), который получен из вируса герпеса, смешали эту ДНК с несколькими миллиграммами ДНК-носителя из спермы лосося и осадили ДНК на культуру L-клеток мыши, в которых ген ТК отсутствовал (ТК-). С частотой 1 на 100000 клетки приобретали ген ТК, поэтому на селективной среде, которая не позволяла расти ТК- - клеткам, росли и нормально размножались ТК+ - клетки.
Другой селективный маркер - ген, кодирующий дигидрофолатредуктазу (ДГФР), можно использовать при трансформации немутантных линий клетки. Благодаря экспрессии многих копий этого гена животная клетка вместе с плазмидой приобретает устойчивость к высоким концентрациям ингибитора фермента, и таким образом трансформантов можно отбирать при высоких концентрациях ингибитора.
Разработано еще два универсальных вектора, содержащих генные маркеры, работающие в нормальных клетках. Они построены по одному и тому же принципу: прокариотические гены, определяющие фенотип трансгенных клеток, соединены с эукариотическими регуляторными сигналами.
Один из векторов состоит из прокариотического гена устойчивости к антибиотику неомицину, встроенного в раннюю область генома SV-40. Эукариотические клетки чувствительны к аналогу неомицина G 418, который инактивируется продуктом гена. Таким образом клетки, прошедшие трасфекцию приобретают способность расти на среде, содержащей G 418.
Генетическая транформация соматических клеток млекопитающих.
Культуры трансформированных клеток млекопитающих используют для получения различных веществ. Хотя культуры клеток животных, особенно при массовом выращивании, гораздо менее экономичны, чем бактериальные дрожжевые культуры, они обладают существенным преимуществом - способностью осуществлять мелкие, но весьма важные модификации белков - продуктов гена млекопитающих. Например, для эффективного функционирования ряда белков необходимо присоединение к ним цепочек из молекул углеводов или липидов. Образование и присоединение таких цепочек - обычный процесс для клеток млекопитающих, тогда как бактериальная клетка не способна производить подобные модификации.
Помимо создания клеток-продуцентов, трансформация соматических клеток млекопитающих позволяет изучать тонкие механизмы регуляции экспрессии генов и целенаправленно модифицировать генетический аппарат клетки животных, а при необходимости и человека, что имеет огромное значение для медицинской генетики.
Генотерапия. Лечение заболеваний с помощью генов получило название генотерапии. Сейчас в мире насчитывается порядка 400 проектов, посвященных лечению с помощью генотеропии.
Разработке программы генной терапии предшествуют тщательный анализ тканеспецифической экспрессии соответствующего гена, идентификация первичного биохимического дефекта, исследование структуры, функции и внутриклеточного распределения его белкового продукта, а также биохимический анализ патологического процесса. Все эти данные учитываются при составлении соответствующего медицинского протокола.
Апробацию процедуры генокоррекции наследственного заболевания проводят на первичных культурах клеток больного, в которых в норме функционально активен данный ген. На этих клеточных моделях оценивают эффективность выбранной системы переноса экзогенной ДНК, определяют экспрессию вводимой генетической конструкции, анализируют ее взаимодействие с геномом клетки, отрабатывают способы коррекции на биохимическом уровне. Используя культуры клеток, можно разработать систему адресной доставки рекомбинантных ДНК, однако проверка надежности работы этой системы может быть осуществлена только на уровне целого организма. Поэтому такое внимание в программах по генной терапии уделяется экспериментам in vivo на естественных или искусственно полученных моделях соответствующих наследственных болезней у животных.
Успешная коррекция генетических дефектов у таких животных и отсутствие нежелательных побочных эффектов генной терапии являются важнейшей предпосылкой для разрешения клинических испытаний.
Таким образом, стандартная схема генокоррекции наследственного дефекта включает серию последовательных этапов. Она начинается созданием полноценно работающей (экспрессирующейся) генетической конструкции, содержащей смысловую (кодирующую белок) и регуляторную части гена. На следующем этапе решается проблема вектора, обеспечивающего эффективную, а по возможности и адресную доставку гена в клетки-мишени. Затем проводится трансфекция (перенос полученной конструкции) в клетки-мишени, оценивается эффективность трансфекции, степень коррегируемости первичного биохимического дефекта в условиях клеточных культур (in vitro) и, что особенно важно, in vivo на животных - биологических моделях. Только после этого можно приступать к программе клинических испытаний.
Существует два типа генотерапии: заместительная и корректирующая.
Заместительная генотерапия заключается во вводе в клетку неповрежденного гена. Внесенная копия заменит по функциям сохранившийся в геноме больного дефектный ген. Все проводимые сегодня клинические испытания используют внесение в клетку дополнительных количеств ДНК.
При корректирующей терапии предполагается замена дефектного гена нормальным в результате рекомбинации. Пока этот метод на стадии лабораторных испытаний, так как эффективность его еще очень низка, но последние исследования показывают успехи в лечении некоторых заболеваний.
Амавроз Лебера - врожденная слепота, редкая форма наследственного заболевания, которое проявляется уже в младенчестве. Из-за дефектного гена (Retinal Pigment Epithelium, 65 kDa) в сетчатке умирают и не восстанавливаются светочувствительные клетки.По статистике, от амавроза Лебера страдает один человек на 81 тысячу. Болезнь сопровождается ослаблением или полной потерей зрения без анатомического нарушения структуры органов.Повреждение гена RPE65 приводит к прекращению синтеза определенных ферментов, участвующих в выработке светочувствительного пигмента, и дегенерации фоторецепторов. Врожденный амавроз Лебера впервые был описан в 1869 году немецким ученым-офтальмологом Теодором Лебером, однако этиология и патогенез этой группы болезней до настоящего времени остаются не до конца изученными.
Клиническими критериями диагностики ВАЛ являются: значительное снижение остроты зрения (от отсутствия реакции на свет и светоощущения до сотых долей), у большинства детей отмечаются плавающие движения глаз, нистагм, окуло-пальцевой симптом, косоглазие, могут встречаться деструкция стекловидного тела и частичное врожденное помутнение хрусталиков. Характерным является резкое снижение скотопических и фотопических показателей суммарного потенциала фоторецепторов сетчатки на электроретинографии (ЭРГ), вплоть до ее отсутствия, при нормальной офтальмоскопической картине глазного дна. Кроме того, отмечаются нарушения цветоощущения от красно-зеленой дисхроматопсии до ахроматопсии, сужение полей зрения до 30-10 градусов, значительное повышение порога электрической чувствительности.
Традиционная лекарственная терапия бессильна в борьбе с этим заболеванием. На помощь пришла генотерапия. Исследователи из США и Англии делали инъекцию вирусного вектора, содержащего исправленный ген в один глаз пациентов, страдающих амаврозом Лебеля. Вектор содержал фермент, необходимый для продукции светочувствительного пигмента и вводился в эпителий пигментного слоя сетчатки. В первом исследовании у всех 12 пациентов светочувствительность в "пролеченном" глазу вернулась. У 4 детей зрение восстановилось до такой степени, что они могли заниматься спортом и нормально учиться в школе. Кроме того, были проведены исследования на саймири (беличтьи обезьянки), страдающих дальтонизмом. Инъекция "исправленных" генов вернула им полное цветовое зрение.
В зависимости от способа введения экзогенных ДНК в геном пациента генная терапия может проводиться либо в культуре клеток (ex vivo), либо непосредственно в организме (in vivo). Клеточная генная терапия или терапия ex vivo предполагает выделение и культивирование специфических типов клеток пациента, введение в них чужеродных генов, отбор трансфецированных клеток и реинфузию их тому же пациенту. Примером может служить лечение комбинированного иммунодефииицита.
Генная терапия in vivo основана на прямом введении клонированных и определенным образом упакованных последовательностей ДНК в специфические ткани больного. В настоящее время не существует общедоступного метода культивирования клеток легких, поэтому при легочных заболеваниях единственный способ доставить чужеродный ген - это ввести его прямо в организм.
Список наследственных заболеваний, которые пытаются или планируют лечить генами, велик. Это и ревматоидный артрит, и фенилкетонурия, и заболевания, связанные с недостатком гормонов (инсулина, эритропоэтина, гормона роста). В случае хронической анемии, связанной с дефицитом эритропоэтина, на основании опытов на животных предлагается принципиально новый подход к лечению. Так как каждая из наших клеток содержит один и тот же геном, можно заставить фибробласты кожи, которые в норме не производят эритропоэтина, синтезировать этот гормон. Для этого нужно ввести в геном новую контролирующую область и тем самым снять запрет со считывания (экспрессии) гена эритропоэтина, присутствующего, но "молчащего" в фибробластах.
Практически в любой области медицины либо начаты клинические испытания лечения наследственных заболеваний с помощью генотерапии, либо в опытах на животных разрабатываются подходы к такому лечению. По мере усовершенствования методов доставки генов и контроля их экспрессии список заболеваний, к которым можно применять генотерапию, будет безусловно расширяться. Огромные перспективы открывает использование генотерапии для лечения онкологических заболеваний. Многолетние усилия ученых привели к пониманию того, что рак — это генетическое заболевание и его развитие происходит многостадийно, в результате серии генетических нарушений, накапливающихся в клетке. Следовательно, каждый из таких отдельных генетических эффектов может стать точкой приложения генотерапевтического подхода.
Генная терапия успешно применяется для лечения не только наследственных, но и значительно более распространенных мультифакториальных болезней (диабет, остеопороз, ревматоидный артрит, различные опухоли). Для лечения таких заболеваний применяется не одна, а сразу много генетических конструкций, исправляющих дефекты различных стадий течения патологического процесса.
Дата добавления: 2016-02-09; просмотров: 2937;