ПРОЦЕСС ТРАНСКРИПЦИИ У ПРО- И ЭУКАРИОТ
Общая характеристика транскрипции. В отличие от репликации ДНК, тесно связанной с клеточным делением, транскрипция ДНК происходит практически во всех ядросодержащих клетках — как делящихся, так и неделящихся.
Причем в делящихся клетках она совершается в любой момент митотического цикла, кроме периода репликации (у эукариот) и собственно деления. У прокариот, видимо, нет и этого ограничения: клеточный цикл бывает столь коротким (20-40 мин), что репликация и транскрипция происходят одновременно, только на разных участках молекулы ДНК (которая у бактерий является кольцевой).
Более того, транскрипция какого-либо участка ДНК может совершаться не только почти в любой момент цикла, но и многократно — сколь угодное число раз. С другой стороны, набор транскрибируемых в клетке участков под действием тех или иных факторов нередко меняется.
Ферментативное обеспечение процесса осуществляется РНК-полимеразой. Фермент ползет вдоль ДНК и катализирует поочередное включение в растущую цепь рибонуклеотидов, комплементарных нуклеотидам матричной цепи ДНК.
Субстратами синтеза РНК являются рибонуклеозидтрифосфаты (рНТФ); как и при синтезе ДНК, в ходе включения в строящуюся цепь они теряют пирофосфатные остатки:
 Свободные остатки рНМФ + пиро
Свободные остатки рНМФ + пиро
рНТФ в строящейся цепи РНК фосфат
Это обеспечивает процесс энергией, так что дополнительных ее источников не требуется.
Еще одно сходство с синтезом ДНК состоит в направлении роста строящейся цепи — 5'→3'. Это значит, что у этой цепи очередные нуклеотиды присоединяются к 3'-концу.
Как при всех матричных синтезах, строящаяся цепь антипараллельна матричной цепи ДНК. Следовательно, последняя транскрибируется ферментом в направлении 3'→5'.
Но имеются и принципиальные отличия от синтеза ДНК.
а) Асимметричность процесса: в качестве матрицы, как мы знаем, используется лишь одна цепь ДНК. Не совсем ясно, как ферментная система осуществляет правильный выбор нужной цепи. Видимо, ключевую роль тут играют какие-то последовательности нуклеотидов на одной из цепей, узнаваемые системой.
б) Консервативность процесса: молекула ДНК по окончании синтеза РНК возвращается в исходное состояние. При синтезе же ДНК молекулы наполовину обновляются, что делает репликацию полуконсервативной.
в) Беззатравность процесса: синтез РНК не требует для своего начала никакой затравки, тогда как при репликации ДНК необходима РНК-затравка.
Процесс транскрипции у прокариот и эукариот существенно различается.
Основные этапы транскрипции:
1. Инициация
2. Элонгация
3. Терминация
4. Процессинг
Особенности транскрипции у прокариот. Осуществляется ДНК-зависимой РНК полимеразой - ферментом катализирующим синтез РНК, который состоит из нескольких субъединиц: двух a, одной b, одной b’ и одной s. Такой комплекс (2abb’s) называется холо-ферментом и имеет молекулярную массу около 500 000 Да. Фермент лишенный s-субъединицы называется кор-ферментом (рис.4.7).
Две a субъединицы представляют собой каркас РНК-полимеразы. К ним крепятся остальные субъединицы. b’- субъединица отвечает за прочное связывание с ДНК за счет кластера положительно заряженных аминокислот. В b- субъединице находятся два каталитических центра. Один отвечает за инициацию, а другой - за элонгацию. Один центр работает в холо-, а другой - в кор- ферменте.
Только холо-фермент обладает высоким сродством к специфической последовательности нуклеотидов - промотору, сродство к остальным случайным последовательностям ДНК у него снижено в 10000 раз. У кор-фермента одинаковое сродство к любой последовательности нуклеотидов.
Сам по себе s- фактор обладает наименьшим сродством к ДНК по сравнению с другими субьединицами РНК-полимеразы, однако он придает холо-ферменту такую конформацию, которая обладает повышенным сродством к промотору.
1. Инициация.
Для инициации транскрипции необходим холофермент, нуклеозидтрифорфат ( всегда АТФ или ГТФ) и наличие специального участка в ДНК называемого промотором.
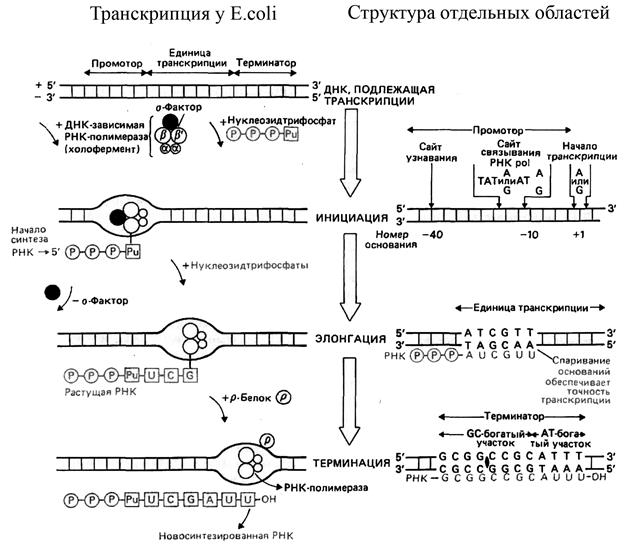
Рис. 4.7. Процесс транскрипции у прокариот
Промотор – это участок молекулы ДНК имеющий размер около 40 пар оснований и расположенный непосредственно перед участком инициации транскрипции. Изучение нуклеотидной последовательности более чем 50 разных промоторных участков ДНК прокариот выявило только два консервативных участка, по-видимому играющих ключевую роль в узнавании и функционировании промотора. Одна из этих последовательностей состоит из шести или семи пар оснований и расположена на расстоянии примерно 10-15 оснований до того нуклеотида, с которого начинается транскрипция (+1). Этот участок обычно обозначают как –10-последовательность, или бокс Прибнова в честь ее открывателя и имеет общий вид ТАТ(Рu)АТ(Рu), где Pu- пурин. Вторая последовательность, длина которой обычно равна девяти нуклеотидам, расположена на расстоянии приблизительно 35 оснований до сайта инициации
(–35-последовательность) и также встречается в большинстве промоторов прокариот. Нуклеотидная последовательность сегмента ДНК между –35- и –10- участками не является критической, важно лишь расстояние между этими участками.
–35-последовательность участвует в связывании РНК-полимеразы, которое предшествует перемещению фермента в бокс Прибнова. Когда полимераза связывается с проморотом, происходит локальное расплетение двойной спирали ДНК на протяжении примерно 1,5 витка (15 нуклеотидных пар) и образуется «открытый промоторный комплекс». Первым в строящуюся цепь РНК всегда включается пуриновый нуклеотид – АТФ или ГТФ, причем все три его фосфатных остатка сохраняются. Затем образуется первая 5’,3’-фосфатная связь со вторым нуклеотидом.
2. Элонгация цепи РНК – это стадия транскрипции, которая наступает после присоединения примерно 8 рибонуклеотидов. В этот момент РНК-полимераза претерпевает структурное изменение при котором от комплекса отделяется s-субъединица и остается кор-фермент, катализирующий дальнейшее удлинение растущей цепи пре-РНК до окончательного размера по мере продвижения РНК-полимеразы по ДНК.
Соответственно, перемещается и «открытый промоторный комплекс», т.е. участок локального расплетения ДНК. Раскрытая ферментом область составляет только несколько пар оснований. На транскрибированной же части ДНК двухцепочечная спиральная структура восстанавливается сразу после ухода РНК-полимеразы.
Примерная скорость движения фермента и синтеза РНК — 30-40 нуклеотидов в секунду.
Транскрипция может сопровождаться ошибками спаривания, в результате которых в синтезируемую РНК включаются «неправильные» нуклеотиды. В среднем одна такая ошибка приходится на 2 х 104 включенных нуклеотидов. Это существенно чаще, чем появление нерепарированных ошибок репликации.
Очевидно, меньшая точность транскрипции связана с тем, что ошибки здесь имеют не столь серьезные последствия. Они легко компенсируются благодаря образованию на одном гене множества копий пре-РНК.
Кроме того, вследствие вырожденности генетического кода, не каждая замена нуклеотида меняет смысл кодона мРНК. Можно найти, что в 67 % ошибок, касающихся третьих положений кодонов, смысл последних остается прежним.
3. Терминация (прекращение роста) цепи РНК происходит на специфических участках ДНК, называемых терминаторами. Начало этих участков обычно обогащено ГЦ-парами, а остальная последовательность АТ-парами. Поскольку сила взаимодействия пар Г-Ц довольно велика, локальное расплетение таких участков ДНК происходит трудней. Это замедляет продвижение РНК-полимеразы и может служить для нее сигналом к прекращению транскрипции.
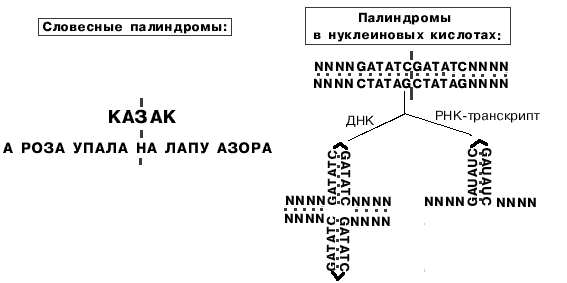
ГЦ-богатый участок ДНК часто представляет собой палиндром.
Палиндромы - последовательности, которые читаются одинаково слева направо и справа налево.
Палиндромы первого порядка имеют одну ось симметрии, второго - две, третьего - три.
Но еще до окончания процесса в конце новосинтезированной РНК тоже успевает появиться ГЦ-богатый участок. Благодаря взаимодействию между своими нуклеотидами, он образует «шпильку». Т. е. взаимодействия с нуклеотидами матричной цепи ДНК заменяются на «внутришпилечные» взаимодействия. Это облегчает отсоединение РНК от ДНК.
У бактерий тому же самому часто способствует и специальный белок —Rho-фактор (ρ-фактор). Он движется по ДНК вслед за РНК-полимеразой, догоняет ее на ГЦ-участке ДНК и, обладая расплетающей активностью, облегчает расхождение цепей РНК и ДНК.
4. Посттранскрипционный процессинг – это процесс созревания при котором первичные пре-РНК модифицируются и превращаются в зрелую РНК. Характер и степень модификации пре-РНК зависит от типа РНК.
У прокариот мРНК не подвергается процессингу. У некоторых бактерий транскрипция и трансляция сопряжены т.е. происходят одновременно: 5’-конец мРНК может транслироваться на рибосоме и затем подвергаться деградации еще до заверщения синтеза ее 3¢-конца.
Молекулы тРНК вначале синтезируются в виде пре-тРНК, которая примерно на 20% длиннее, чем соответствующая тРНК. Лишние последовательности, расположенные у 5¢ и 3¢-концов удаляются с помощью таких ферментов как рибунуклеазы Q и Р. Иногда молекула пре-тРНК состоит из двух или более молекул тРНК соединенных между собой. Их разделение также осущетвляется также рибонуклеазами.
Гены рРНК прокариот расположены в транскрипционных блоках. Например три гена рРНК кишечной палочки располагаются вместе с генами нескольких тРНК в одном таком блоке и транскрибируются в виде одной молекулы РНК. Эти молекулы рРНК и тРНК отделены друг от друга спейсерной РНК. Расщепление первичного транскрипта на отдельные составляющие катализирует рибонуклеаза Q.
Особенности транскрипции у эукариот. 1. У эукариот для транскрипции используются три ДНК-зависимых РНК-полимераз. РНК-полимераза I локализована в ядрышке, где она катализирует синтез пре-рРНК в виде большого первичного транскрипта содержащего молекулы рРНК 18S 5,8S и 28S. РНК-полимераза II находится в нуклеоплазме и участвует в синтезе первичного транскрипта мРНК. РНК-полимераза III также локализована в нуклеоплазме и участвует в синтезе тРНК и 5S-рРНК (малая рРНК).
2. В случае эукариот промотор – более сложное понятие, поскольку РНК-полимераза связывается с ДНК не непосредственно, а лишь вместе с комплексом других белков – т.н. общих факторов транскрипции.
Наиболее часто (у 80% промоторов эукариот) различают небольшую область инициации, ТАТА-бокс (сходный с боксом Прибнова у бактерий) и ряд других типичных участков – ГЦ-, ЦААТ- и другие боксы. Очередность их расположения в разных промоторах неодинакова: в одних случаях ГЦ-бокс предшествует ТАТА-боксу, а в других – наоборот, следует после него.
3. На этапе инициации транскрипции у эукариот всегда требуется предварительное связывание с промотором целой совокупности белков – общих факторов транскрипции, с образованием комплекса TFIID (Transcriptional Factor D for polymerase II). Из обозначения следует, что эти белки необходимы для связывания с промотором РНК-полимеразы II. Если же ген транскрибируется РНК-полимеразой I или III, его промотор содержит вместо ТАТА-бокса какую- то иную последовательность и используются другие факторы транскрипции.
4. У эукариот более сложными являются и сигналы терминации. В отличие от бактериальных, они тоже транскрибируются РНК-полимеразой, и только затем последняя завершает свою работу.
5. В результате транскрипции у эукариот (в отличие от прокариот) образуются лишь предшественники тех или иных РНК — мРНК, рРНК и тРНК.
В строении зрелых цепей РНК каждого вида и соответствующих пре-РНК существуют следующие различия:
а) Пре-мРНК (рис.4.8). Эти цепи обычно в несколько раз длиннее, чем зрелые мРНК. Они, во-первых, включают транскрипты спейсеров (представляющих собой регуляторные участки, отделы со структурной ролью и т. д.). Во-вторых, кодирующая часть пре-мРНК, как и в исходном гене, прерывается интронами. При этом интронные последовательности нередко образуют «шпильки».
Длина пре-мРНК не только гораздо больше, чем у мРНК, но и значительно сильней варьирует у разных молекул — от 2 тыс. до 20 тыс. нуклеотидов. Поэтому данный вид РНК часто называют гетерогенной ядерной РНК (гяРНК).
Другая особенность пре-мРНК — отсутствие на 5'-конце «колпачка» (кэпа), а на 3'-конце — поли(А)-фрагмента.
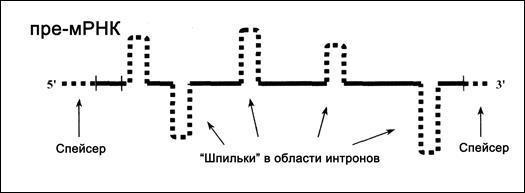
Рис. 4.8. Продукт транскрипции – пре-мРНК
(«лишние» нуклеотидные последовательности показаны пунктиром)
В большинстве случаев пре-мРНК эукариот несут информацию о синтезе лишь одной пептидной цепи, т. е. дают при созревании только одну молекулу мРНК.
Среди исключений из этого правила — гистоновая пре-мРНК. Кластер гистоновых генов транскрибируется как единое целое, а при созревании пре-мРНК из последней «нарезаются» 5 разных гистоновых мРНК.
Наличие подобных исключений — еще одно отличие пре-мРНК от мРНК: все зрелые мРНК эукариот, без каких-либо исключений, являются моноцистронными.
б) Пре-рРНК (рис.4.9). Кластер трех генов рРНК тоже транскрибируется как единое целое. Образующаяся пре-рРНК, или 45S-РНК, содержит последовательности сразу трех зрелых рРНК — 18S-, 5,8S- и 28S-рРНК.
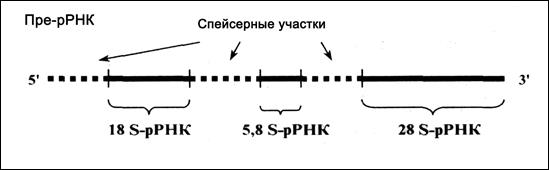
Рис. 4.9. Продукт транскрипции – пре-рРНК
(«лишние» нуклеотидные последовательности показаны пунктиром)
Эти последовательности разделены спейсерами, но не содержат интронов. Кроме того, в них нет модифицированных нуклеотидов, содержащихся в зрелых рРНК.
в) Пре-тРНК (рис.4.10). В отличие от пре-рРНК, все пре-тРНК содержат последовательности лишь одной тРНК.
При этом уже на уровне пре-тРНК образуется типичная структура «клеверного листа». Однако последняя еще отличается от окончательной: в ней
 - имеется ряд дополнительных последовательностей (с обеих концов и в середине молекулы),
- имеется ряд дополнительных последовательностей (с обеих концов и в середине молекулы),
- отсутствуют минорные нуклеотиды,
- не сформирована типичная последовательность акцепторной петли (ЦЦА),
- антикодон не занимает своего «правильного» положения.
Из всего вышесказанного ясно, что непосредственные продукты транскрипции действительно должны подвергаться определенным (и существенным) изменениям для того, чтобы превратиться в функционально активные цепи РНК.
Созревание (процессинг) РНК. Практически все процессы созревания РНК могут быть подразделены на три типа:
· удаление одних,
· присоединение других и
· модификация тех же или третьих нуклеотидов.
Удаление «лишних» нуклеотидов осуществляется специальными нуклеазами. Экзонуклеазы последовательно отщепляют с определенного конца цепи (3' или 5') по одному нуклеотиду. А эндонуклеазы разрезают цепь где-то в средних участках, приводя к ее фрагментации.
Процессы созревания, идущие с участием нуклеаз:
1. Отщепление отдельных «лишних» нуклеотидов с концов цепи. Так, в пре-РНК с 5'-конца всегда находится АТФ или ГТФ, а с 3'-конца нередко — ГЦ-участки. Они играют важную роль в самом процессе транскрипции, но не нужны и даже мешали бы при функционировании зрелой цепи, и поэтому удаляются.
2. С концов цепей отщепляются спейсерные последовательности нуклеотидов (если таковые имеются). Обычно для этого используются эндонуклеазы.
3. Такие пре-РНК, как 45S-пре-рРНК и гистоновая пре-мРНК, разрезаются эндонуклеазами на индивидуальные цепи РНК.
4. Из средних участков пре-тРНК и практически всех (кроме гистоновых) пре-мРНК вырезаются интронные последовательности. При этом экзонные последовательности сшиваются друг с другом в непрерывную цепь. Данный процесс (вырезание интронов и сшивание экзонов) обозначается как сплайсинг.
Очевидно, для его осуществления требуются не только эндонуклеазы, но и лигазы (катализирующие соединение экзонов).
Механизм сплайсинга
Один из ключевых моментов механизма сплайсинга (рис.4.11) — обеспечение точности разрезания цепи пре-РНК: ошибка даже на один нуклеотид приведет к «сдвигу рамки», что изменит смысл всех кодонов мРНК или антикодона тРНК.
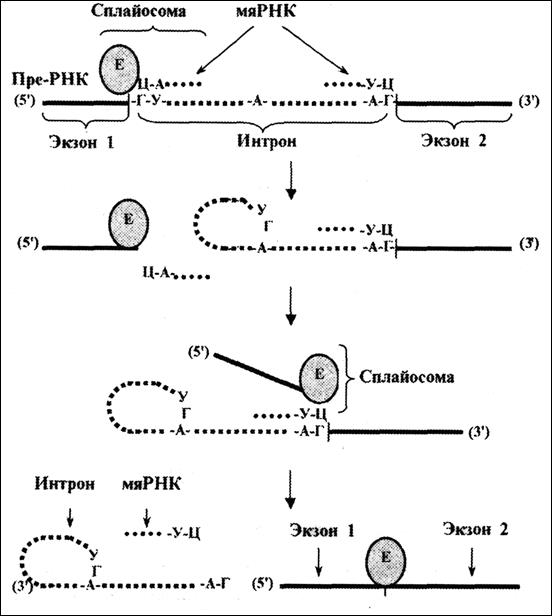
Рис. 4.11. Механизм сплайсинга.
Е-ферментный комплекс (с нуклеазной и лигазной активностью)
Точность разрезания достигается благодаря двум обстоятельствам. Во-первых, в начале и в конце каждого интрона имеются определенные последовательности нуклеотидов: так, интроны всегда начинаются с Г-У, а кончаются дуплетом А-Г. Во-вторых, для узнавания этих последовательностей используются специальные РНК — т. н. малые ядерные РНК (мяРНК). Последние связаны с ферментами, катализирующими сплайсинг. Такие рибонуклеопротеидные комплексы называются сплайосомами.
Сплайсинг начинается со взаимодействия двух мяРНК с началом и концом интрона. Это дает «ориентацию» для эндонуклеазы: последняя действует на границах двух- и одноцепочечных участков.
Первый разрыв пре-РНК происходит в области 5'-конца интрона — на рис. 4.11. это место нахождения левого края левой мяРНК. При этом 5'-конец интрона связывается с одним из нуклеотидов в средней части того же интрона, что приводит к образованию кольцевой (или, более точно, лассоподобной) структуры.
Поэтому первая (на рисунке — левая) мяРНК, видимо, диссоциирует, а ферментный комплекс перемещается к другой (на рисунке — правой) мяРНК, маркирующей 3'-конец интрона. Здесь происходит второй разрыв пре-РНК — по месту нахождения «правого» конца «правой» мяРНК. Точнее, связь экзона 2 с интроном заменяется на связь с экзоном 1.
Следовательно, оба разрыва совершаются одним и тем же способом — путем замещения одной межнуклеотидной связи на другую такую же связь.
Присоединение и модификация нуклеотидов
В процессе созревания пре-РНК происходит также и нетранскрипционное присоединение отдельных нуклеотидов.
1. В случае пре-мРНК со стороны 5'-конца присоединяется (с помощью нетипичной для полинуклеотидов пирофосфатной связи) 7-метилгуаниловый нуклеотид — компонент «колпачка». А со стороны 3'-конца понуклеотидно наращивается поли(А)-фрагмент примерно из 200 нуклеотидов. Для этого используются специальные ферменты; в частности, для образования поли(А)-фрагмента — полиаденилатполимераза.
2. В случае пре-тРНК с 3'-конца по очереди присоединяются три нуклеотида — Ц, Ц и А, образующие акцепторную ветвь.
Наконец, важный момент созревания пре-РНК — образование в их составе модифицированных нуклеотидов. Как и в случае метилирования ДНК, минорные нуклеотиды появляются в полинуклеотидной цепи не на стадии полимеризации, а по завершении ее — путем модификации содержащихся в цепи обычных нуклеотидов.
Так, в пре-мРНК происходит метилирование рибозных остатков нуклеотидов «колпачка», а в пре-рРНК — тоже метилирование рибозных остатков, но содержащие их нуклеотиды расположены по всей длине цепи — с частотой примерно 1 %.
Гораздо более многообразны процессы модификации в случае пре-тРНК. Например, определенные остатки уридина подвергаются восстановлению (с образованием дигидроуридина), другие — изомеризации (что дает псевдоуридин), третьи — метилированию (метилуридин). Некоторые остатки аденозина дезаминируются (превращаясь в инозин), причем часть из продуктов затем еще метилируется (образуя метилинозин). И так далее: список происходящих модификаций в пре-тРНК можно продолжить.
Все вышеперечисленные события и приводят в конце концов к образованию в ядре зрелых молекул РНК:
а) 4 –х видов рРНК: 28S-, 18S-, 5,8S- и 5S-РНК;
б) нескольких десятков видов тРНК — по 1-3 (или даже больше) для каждой из 20 аминокислот;
в) тысяч различных мРНК — копий генов, функционирующих в данных клетках.
рРНК и мРНК тут же, в ядре, связываются с белками. При этом рРНК формируют с рибосомными белками большие и малые субъединицы рибосом, которые затем выходят в цитоплазму. А мРНК связываются с другими белками (выполняющими, видимо, защитную и транспортную функции) и в комплексе с ними тоже перемещаются в цитоплазму.
В процессе всех этих преобразований существенно меняются седиментационные свойства. Для иллюстрации приведем стадии созревания одной из мРНК (табл. 4.1).
Таблица 4.1. Созревание мРНК
| Последовательные формы | 40 S-пре-мРНК | 15 S-пре- мРНК |
16,5 S-поли(А)- мРНК |
27 S-поли(А)- мРНП |
| Кол-во нуклеотидов в РНК | ~9200 | ~1200 | ~1430 | ~1430 |
| Кол-во связанных молекул белка | — | — | — | 3 (с массой по 50.000) |
Как видно, в результате удаления «лишних» нуклеотидов цепь пре-мРНК укорачивается примерно на 87 % (т. е. остается лишь 13 % от исходной длины). Затем присоединяются примерно 230 адениловых нуклеотидов, отчего коэффициент седиментации несколько увеличивается. И, наконец, на последней стадии он возрастает гораздо сильней — в результате связывания с мРНК трех белковых молекул. В итоге образуется мРНП — м-рибонуклеопротеид, т. е. комплекс мРНК с белками.
Конвейерный характер процесса. До сих пор мы имели в виду одну молекулу РНК-полимеразы и образование одной цепи пре-РНК.
Но в действительности через какое-то время после того, как предыдущая молекула РНК-полимеразы, покинув промотор, продвинется по ДНК на некоторое расстояние, с промотором связывается следующая молекула фермента и тоже начинает транскрипцию.
Поэтому на каждом транскрибируемом гене обычно работают, двигаясь друг за другом, сразу несколько молекул РНК-полимеразы (рис. 4.12).
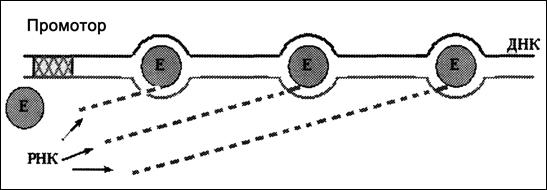
Рис. 4.12. Транскрипция ДНК одновременно несколькими молекулами
РНК-полимеразы (Е)
Среднее расстояние между ними зависит от «силы» промотора (во многом обусловленной транскрипционными факторами) и концентрации РНК-полимеразы. Примерный порядок этого расстояния — 300-500 н. п.
Соответственно, с одним геном одновременно связано несколько растущих цепей пре-РНК.
Таким образом, транскрипция гена происходит конвейерным способом.
Дата добавления: 2016-02-09; просмотров: 5894;