ТРАНСКРИПЦИЯ ДНК. СТРУКТУРА МОЛЕКУЛЫ РНК
Все транскрипционные факторы, как и сама транскрипция, призваны обеспечить только одно — образование с нужной скоростью РНК на тех или иных участках хромосом.
Прежде чем говорить о механизме данного процесса, кратко охарактеризуем строение разных видов РНК. Это необходимо, чтобы понять особенности транскрипции и феномен созревания новосинтезированных молекул РНК.
Содержание РНК в любых клетках в 5 –10 раз превышает содержание ДНК. Основная роль РНК состоит в трансляции генетической информации с образованием белков. Однако молекулы РНК принимают участие и в осуществлении некоторых специализированных эндонуклеазных функций, возможно регулирующих различные этапы экспрессии генов. Молекулами РНК представлены геномы некоторых вирусов (ретровирусов и множества вирусов животных, растений и насекомых с одно-и двухцепочечным геномом). Во всех клетках присутствуют следующие виды РНК: рибосомная РНК (рРНК), транспортная РНК (тРНК) и информационная, или матричная, РНК (мРНК). Большинство (если не все) клеток содержат также много других малых цитоплазматических РНК (мцРНК),а в клетках эукариот присутствует еще и множество малых ядерных РНК (мяРНК).Около 80% массы клеточных РНК составляют три или четыре вида рРНК, а около 15%– почти 100 видов тРНК. На долю нескольких тысяч различных матричных РНК приходится менее 5%клеточной РНК, а на долю малых ядерной и цитоплазматической РНК, число видов которых пока неизвестно,– менее 2%от общего количества.
4.1.1. Общий план строения РНК
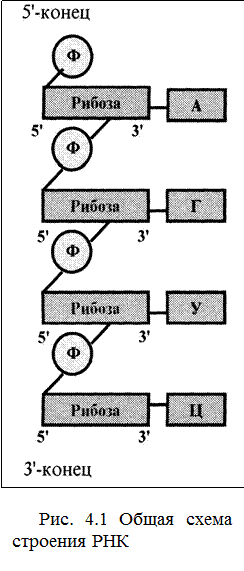
Как и ДНК, РНК представляют собой линейные (т. е. неразветвленные) полинуклеотиды с тем же принципом организации (рис.4.1):
· состоят из четырех видов нуклеотидов, каждое из которых включает азотистое основание, пентозу и фосфатный остаток;
· нуклеотиды связаны в цепь с помощью 5',3'-фоcфодиэфирных связей;
· полинуклеотидные цепи полярны, т. е. имеют различимые 5'- и 3'-концы.
Но имеются и отличия от ДНК. Главное из них — то, что молекулы РНК (кроме РНК некоторых вирусов) являются не двух-, а одноцепочечными. Причиной служат следующие три особенности первичной структуры.
а) Во-первых, пентоза в РНК — это не дезоксирибоза, а рибоза, которая содержит дополнительную гидроксигруппу. Последняя делает двухцепочечную структуру менее компактной.
б) Во-вторых, среди четырех главных, или мажорных, азотистых оснований вместо тимина содержится урацил, отличающийся от тимина лишь отсутствием метильной группы в 5-м положении.
Благодаря этому уменьшается сила гидрофобного взаимодействия в комплементарной паре А-У. Что тоже снижает вероятность образования устойчивых двухцепочеч ных молекул.
в) Наконец, в РНК (особенно в тРНК) высоко содержание т. н. минорных оснований и нуклеозидов. Среди них — дигидроуридин (в урациле нет одной двойной связи), псевдоуридин (урацил иначе, чем обычно, связан с рибозой), диметиладенин и диметилгуанин (в азотистых основаниях — по две дополнительных метильных группы) и многие другие.
Почти все эти основания не могут участвовать в комплементарных взаимодействиях. Так, метильные группы в диметиладенине (в отличие от тимина и 5-метилцитозина) находятся при таком атоме, который в паре А-У образует водородную связь; следовательно, теперь данная связь замкнуться не может. Это тоже препятствует образованию двухцепочечных молекул.
Таким образом, широко известные отличия состава РНК от ДНК имеют огромное биологическое значение: ведь свою функцию молекулы РНК способны выполнять только в одноцепочечном состоянии. Наиболее очевидно это для мРНК: трудно представить, как бы могла двухцепочечная молекула транслироваться на рибосомах.
Вместе с тем, оставаясь одиночной, в некоторых участках цепь РНК может образовывать петли, или «шпильки», с двухцепочечной структурой
Эта структура стабилизирована взаимодействием оснований в парах А:::У и Г:::Ц. Однако могут образовываться и «неправильные» пары (например, Г····У), а в некоторых местах «шпильки» и вообще не происходит никакого взаимодействия.
В составе таких петель может содержаться (особенно в тРНК и рРНК) до 50 % всех нуклеотидов.
Общее же содержание нуклеотидов в РНК варьирует от 75 единиц до многих тысяч. Но даже самые крупные РНК на несколько порядков короче хромосомных ДНК.
Теперь обратимся к особенностям структуры трех классов РНК — мРНК, тРНК и рРНК.
Особенности строения мРНК
В начале 50-х годов Ф.Крик сформулировал свою знаменитую центральную догму молекулярной биологии, согласно которой генетическая информация от ДНК к белкам передается через РНК по схеме ДНК→РНК→белок. Процесс синтеза РНК на матрице ДНК называется транскрипцией, а процесс синтеза белка на матрице РНК – трансляцией. В 1956-1957 годах А.Н.Белозерский и А.С.Спирин показали, что при существенных различиях в нуклеотидном составе ДНК из разных организмов нуклеотидный состав суммарных РНК весьма сходен. На основании этих данных они пришли к сенсационному заключению о том, что суммарная РНК клетки не может выступать в качестве переносчика генетической информации от ДНК к белкам, поскольку не соответствует ей по своему составу. Вместе с тем они заметили, что при значительном изменении нуклеотидного состава ДНК при переходе от организма к организму наблюдается некоторый небольшой сдвиг нуклеотидного состава РНК в ту же сторону. Это позволило предположить существование минорной фракции РНК, которая полностью соответствует по своему нуклеотидному составу ДНК и которая может быть истинным переносчиком генетической информации от ДНК к белкам. Целенаправленный поиск такой РНК, предпринятый сразу в нескольких лабораториях мира, увенчался успехом в 1961 году. В том году С.Бреннер, Ф.Жакоб и М.Месельсон, с одной стороны, и Ф.Гро и Дж.Уотнос с сотрудниками – с другой, обнаружили ДНК-подобную РНК у бактерий. В течение последующих двух-трех лет аналогичная РНК была найдена в самых разных эукариотических организмах. Для ее обозначения был предложен термин «информационная, или матричная, РНК (мРНК)».
Поскольку каждая мРНК содержит информацию о составе той или иной полипептидной цепи, количество разных мРНК в клетке очень велико.
Несмотря на это, имеют место два обстоятельства. Во-первых, все эти мРНК составляют лишь небольшую часть общей массы РНК в клетке — около 5 %. Во-вторых, при всем своем многообразии, зрелые мРНК имеют сходный план строения. Он состоит в том, что линейная цепь мРНК содержит несколько областей с различной функциональной ролью (рис. 4.2.).
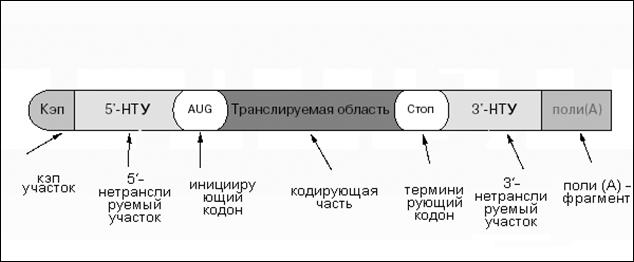
Рис. 4.2. Составные части зрелой мРНК
а) На 5'-конце находится т. н. «колпачок», или кэп — участок из одного-четырех модифицированных нуклеотидов, например:
(7-метил-Г)— ф-ф-ф-(2-О'-метил-Х)-ф-(2-О'-метил-У)-ф--
Первым всегда идет 7-метилгуанилат. Причем с очередным нуклеотидом он связан пирофосфатной связью. Несколько следующих нуклеотидов могут быть метилированы по 2'-положению рибозы. Такая необычная структура призвана защищать 5'-конец мРНК от экзонуклеаз.
б) За «колпачком» идет 5'-нетранслируемый участок — последовательность из нескольких десятков нуклеотидов. Она комплементарна одному из отделов той рРНК, которая входит в малую субъединицу рибосомы. За счет этого она служит для первичного связывания мРНК с рибосомой, но сама, как видно из названия, не транслируется.
в) Трансляция же (считывание) мРНК начинается всегда с т. н. инициирующего кодона. Во всех мРНК он всегда один и тот же — АУГ, т. е. кодирует метионин. Поэтому после синтеза пептидной цепи с ее N-конца, как правило, отщепляется метионин (если последний не нужен для функционирования белка).
г) За инициирующим кодоном в мРНК следует кодирующая часть, которая, собственно, и содержит информацию о последовательности аминокислот в белке.
У эукариот зрелые мРНК являются моноцистронными, т. е. каждая из них несет информацию о структуре только одной полипептидной цепи.
Другое дело, что иногда пептидная цепь вскоре после образования на рибосоме разрезается на несколько более мелких цепей. Так бывает, например, при синтезе инсулина и целого ряда олигопептидных гормонов.
В отличие от эукариот, у бактерий мРНК — полицистронные, и в таком виде они транслируются рибосомами: на разных цистронах мРНК одновременно синтезируются разные полипептидные цепи.
И у прокариот, и у эукариот кодирующая часть зрелой мРНК лишена интронов — каких-либо вставочных некодирующих последовательностей. Иными словами, имеется непрерывная последовательность смысловых кодонов, которая должна читаться в направлении 5'→3'.
д) По окончании этой последовательности находится кодон терминации — один из трех «бессмысленных» кодонов: УАА, УАГ или УГА .
е) За этим кодоном может следовать еще 3'-нетранслируемый участок, значительно превышающий по длине 5'-нетранслируемую область.
ж) Наконец, почти все зрелые мРНК эукариот (кроме гистоновых мРНК) на 3'-конце содержат поли(А)-фрагмент из 150-200 адениловых нуклеотидов.
И 3'-нетранслируемый участок, и поли(А)-фрагмент имеют отношение к регуляции продолжительности жизни мРНК, поскольку разрушение мРНК осуществляется 3'-экзонуклеазами.
Поли(А)-фрагмент отчасти подобен по функции теломере ДНК. В соответствии с гипотезой «билетиков», после того, как очередная рибосома заканчивает трансляцию мРНК, от поли (А)-фрагмента отщепляются 10-15 нуклеотидов. Когда данный фрагмент исчерпывается, начинает разрушаться значащая часть мРНК (если отсутствует 3'-нетранслируемый участок).
Общее количество нуклеотидов в мРНК обычно варьирует в пределах нескольких тысяч. При этом на кодирующую часть иногда может приходиться лишь 60—70 % нуклеотидов.
В клетках молекулы мРНК практически всегда связаны с белками. Последние, вероятно, стабилизируют линейную структуру мРНК, т. е. предупреждают образование в кодирующей части «шпилек». Кроме того, белки могут защищать мРНК от преждевременного разрушения. Такие комплексы мРНК с белками иногда называют информосомами.
Особенности строения тРНК. Основное назначение транспортной РНК (тРНК) – доставлять активированные остатки аминокислот в рибосому и обеспечивать их включение в синтезирующуюся белковую цепь в соответствии с программой, записанной генетическим кодом в матричной РНК.
Удивительна история открытия тРНК. В 50-х годах стало ясно, что информация для аминокислотной последовательности белков закодирована в виде нуклеотидной последовательности матрицы. Хотя генетический код еще не был раскрыт, Г.Гамов предложил считать, что он должен быть триплетным, то есть каждая аминокислота должна кодироваться тройкой нуклеотидов матрицы. Между тем триплетный кодон не может непосредственно узнавать боковые радикалы аминокислот, поскольку между ними нет стерического соответствия. Исходя из этого, Ф. Крик в 1955 году предположил, что должно существовать семейство малых молекул РНК с двойной функцией – каждая из них должна ковалентно связываться с определенным аминокислотным остатком и в то же время иметь в своем составе нуклеотидный триплет, комплементарный кодону для этой аминокислоты. Эти гипотетические РНК должны выполнять функцию адапторов (переходников) между аминокислотой и соответствующим ей кодоном, в связи с чем гипотеза получила название адапторной. Поскольку аминокислот 20, то и соответствующих им РНК-адапторов должно быть не меньше. Ф Крик предположил, что их ковалентное связывание осуществляет 20 специальных ферментов, по одному на каждую аминокислоту. В течение последующих двух лет адапторная гипотеза была доказана экспериментально: были открыты сравнительно небольшие тРНК и специфические ферменты, присоединяющие к ним аминокислотные (аминоацильные) остатки. Далее было показано, что образующиеся аминоацил-тРНК способны переносить остатки аминокислот в растущую цепь белка. После открытия мРНК стало ясно, что действительно молекулы тРНК играют ключевую роль в экспрессии генов, обеспечивая перевод информации, записанной в м РНК в виде последовательности кодонов, на язык аминокислотной последовательности белков. Так, сложнейший биологический процесс был предсказан до того, как обнаружили участвующие в нем макромолекулы.
Первичная, вторичная и третичная структуры. Количество различных тРНК — несколько десятков: от одного до шести видов для каждой из 20 аминокислот. Виды тРНК, способные связывать одну и ту же аминокислоту, называются изоакцепторными. Специфичность тРНК обозначается верхним индексом, например: тРНКАла.
Общее число нуклеотидов в молекуле тРНК — невелико: в среднем от 74 до 95 аминокислотных остатков. Среди них высоко содержание минорных, или модифицированных, нуклеотидов. Например, в аланиновой тРНК таковыми являются 13 % нуклеотидов. В их составе — следующие «нетипичные» нуклеозиды:
— дигидроуридин (дгУ) и псевдоуридин (обозначается y)
— инозин (И): по сравнению с аденозином, аминогруппа замещена на кетогруппу;
— метилинозин (мИ), метил- и диметилгуанозин (мГ и м2Г);
— метилуридин (мУ): то же самое, что риботимидин.
Общий список выявленных в тРНК модифицированных нуклеозидов превышает 60 названий.
Другая особенность состоит в том, что, благодаря образованию нескольких «шпилек», цепь тРНК всегда приобретает характерную структуру «клеверного листа». «Клеверный лист»-это структурное представление молекулы тРНК, полученное из условия образования максимального числа уотсон-криковских пар оснований при данной нуклеотидной последовательности. Участки, в которых с помощью водородных связей образовались такие пары оснований, называются стеблями, а одноцепочечные участки-петлями. Все известные тРНК образуют «клеверный лист» с четырьмя стеблями (акцепторным, D, антикодоновым и Т) и тремя петлями (D, антикодоновой и Т). Некоторые тРНК имеют дополнительные петли и/или стебли (например, вариабельная петля - V-петля дрожжевой тРНК).
Поскольку минорные нуклеотиды, как правило, не способны к .комплементарным взаимодействиям, они содержатся, в основном, в одноцепочечных локусах.
Для одноцепочечных участков приняты следующие названия:
Антикодоновая петля — участок из 7 нуклеотидов в середине цепи. Три из этих нуклеотидов выполняют функцию антикодона, который комплементарно взаимодействует с соответствующим кодовом в цепи мРНК.
Дигидроуридиловая (D-петля), псевдоуридиловая (Т-петля) и имеющаяся не всегда добавочная петли: способствуют формированию специфичной для данной тРНК третичной структуры.
Термин «вторичная структура» относится к расположению в пространстве атомов внутри стебля.
Вторичная структура (рис.4.3) .
Каждый стебель состоит из двух антипараллельных цепей, основания которых образуют друг с другом уотсон-криковские пары с помощью водородных связей. Такое пространственное расположение, установленное экспериментально, было правильно предсказано исходя из структуры клеверного листа. Стебли имеют форму правой двойной спирали, известной как А-форма РНК.
Двойная спираль РНК в А-форме содержит 11 пар оснований на виток, шаг спирали равен 3,1 нм. Расстояние между соседними парами оснований вдоль оси спирали составляет 0,28 нм (3,1 нм/11); они повернуты друг относительно друга на угол 33° (360°/11). Плоскости оснований составляют ~ 20° с нормалью к оси двойной спирали. А-форма РНК близка к А-форме ДНК. РНК не способна переходить в В-форму. Это обусловлено присутствием в ней объемной 2'-гидроксильной группы рибозы, которой нет в дезоксирибозе.
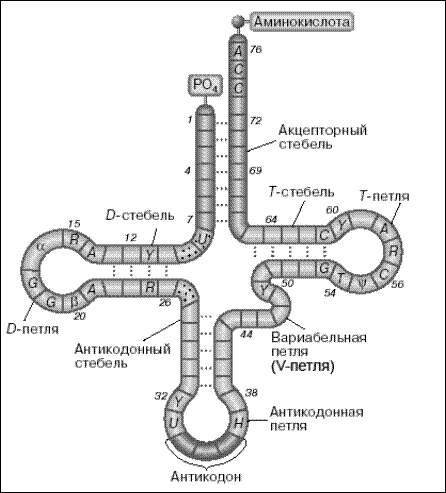
Рис.4.3. Вторичная структура тРНК
Третичная структура - это пространственное расположение всех частей молекулы тРНК.
Третичная структура молекулы напоминает по форме букву Г (рис.4.4). Она значительно более вытянута, чем глобулярные белки той же молекулярной массы. Акцепторный и Т-стебли уложены в пространстве таким образом, что образуют одну непрерывную спираль-«перекладину» буквы Г; антикодоновый и D-стебли образуют «ножку». Эти две части содержат каждая около 10 пар оснований. Почти все основания в тРНК участвуют в ван-дерваальсовых и гидрофобных взаимодействиях, стабилизирующих пространственную структуру молекулы.
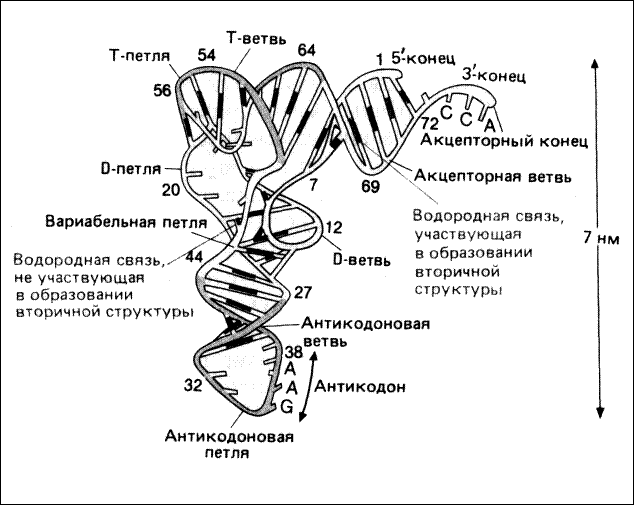
Рис.4.4. Третичная структура тРНК
В тРНК имеются разнообразные водородные связи. В стеблях, являющихся двойными спиралями, реализуются уотсон-криковские водородные связи. Многие основания, расположенные вне стеблей, образуют водородные связи с другими основаниями, но эти связи отличаются от уотсон-криковских. Помимо этого 2'-гидроксильные группы полинуклеотидной цепи также образуют водородные связи с основаниями и с атомами водорода основной цепи.
Взаимодействия тРНК с лигандами
Связывание «своей» аминокислоты c тРНК происходит с помощью фермента — специфической аминоацил-тРНК-синтетазы. Существует 20 видов таких ферментов — по одному на аминокислоту. И в каждом случае фермент имеет 2 центра узнавания — аминокислоты и любой из изоакцепторных тРНК, специфичных для данной аминокислоты.
Следовательно, именно данные ферменты сопрягают генетический код со структурой аминокислот.
Причем в составе ферментов есть не только центр образования ковалентной связи между аминокислотой и тРНК, но и центр гидролиза такой связи. Последний срабатывает, если к тРНК присоединилась «не та» аминокислота.
Что касается взаимодействия тРНК с кодоном мРНК, то оно удовлетворяет общим принципам комплементарности антипараллельности. Последнее означает: поскольку смысл кодона мРНК читается в направлении 5'®3', то антикодон в тРНК должен читаться в направлении 3'→5'.
При этом первые два основания кодона и антикодона спариваются строго комплементарно, т. е. образуются только пары А-У и Г-Ц. Спаривание же третьих оснований может отступать от этого принципа. Допустимые пары определяются схемой:
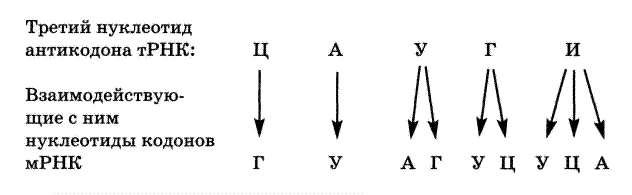
Из схемы вытекает следующее.
а) Молекула тРНК связывается только с 1-м типом кодона, если третий нуклеотид в ее антикодоне — Ц или А.
б) тРНК связывается с 2-мя типами кодонов, если антикодон заканчивается на У или Г.
в) И, наконец, тРНК связывается с 3-мя типами кодонов, если антикодон кончается на И (инозиновый нуклеотид); такая ситуация, в частности, в аланиновой тРНК.
Отсюда, в свою очередь, следует, что для узнавания 61 смыслового кодона требуется, в принципе, не такое же, а меньшее количество разных тРНК.
Особенности строения рРНК. Образование рибосомальной РНК (рРНК) происходит в ядрышке. Эта молекула состоит из 3000 -5000 нуклеотидов. В цитоплазме с молекулами РНК объединяются рибосомальные белки и образуется малая и большая субъединицы рибосом (рис.4.6).
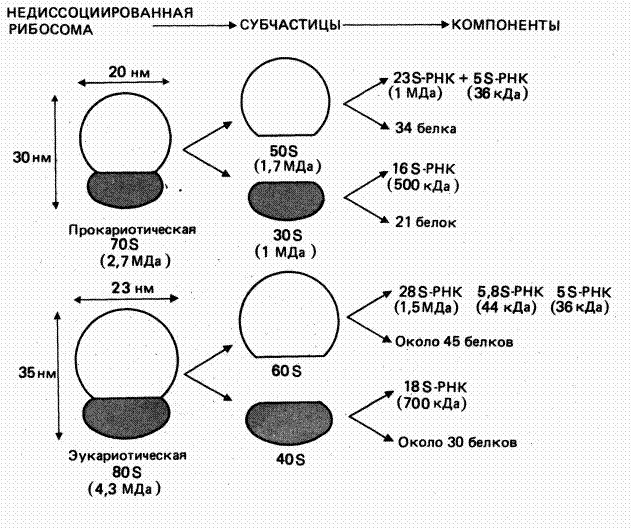
Рис.4.6. Структура рибосом
От общей массы РНК клетки на долю рРНК приходится до 90%.
Субъединицы рибосом, а также входящие в них рРНК принято обозначать по их константе седиментации*.
В результате каждая субъединица рибосомы — это свернутый рибонуклеопротеидный тяж.
Среди азотистых оснований в рРНК выше, чем обычно, содержание гуанина и цитозина. Поэтому и гены рРНК тоже являются ГЦ-богатыми .
Дата добавления: 2016-02-09; просмотров: 1636;