МОЛЕКУЛЯРНЫЙ МЕХАНИЗМ РЕПЛИКАЦИИ ДНК
Один из важнейших внутриядерных процессов – репликация, или удвоение количества ДНК. Именно репликация ДНК обеспечивает воспроизведение наследственной информации при образовании новых клеток. Примерно в то же время происходит удвоение количества и хромосомных белков. Следовательно, репликация ДНК – составная часть более сложного процесса – репликации хромосом.
Репликативный синтез ДНК тесно связан с делением клетки.
Как известно, клеточные деления бывают двух типов: мейоз и митоз. Мейоз используется лишь в одном случае: так проходит последнее деление предшественников половых клеток. Остальные деления осуществляются путем митоза; это все предыдущие деления предшественников половых клеток, а также все деления соматических клеток.
Если обозначить количество ДНК в одинарном наборе хромосом через n, то его изменение в ходе указанных двух типов деления отразится рис. 2.1.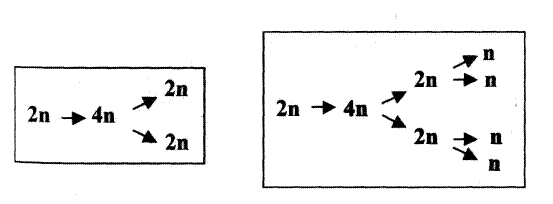
Рис. 2.1. Схемы митоза (слева) и мейоза (справа)
В обоих случаях ядро исходной клетки содержит обычный двойной набор хромосом и в нем – диплоидное (2n) количество ДНК; при подготовке к делению это количество путем репликации возрастает до тетраплоидного (4n), а в ходе последующего деления распределяется поровну между двумя дочерними клетками.
Но при мейозе добавляется еще одно деление, причем без предшествующей репликации ДНК; так что в итоге образуются четыре клетки с одинарным, или гаплоидным (п), количеством ДНК.
Следовательно, среди всех делений эукариотических клеток лишь в одном случае (во втором делении мейоза) не происходит предшествующего удвоения ДНК.
Отличным является и тот период клеточного цикла, когда совершается репликация ДНК.
В мейотических клетках (сперматоцитах и ооцитах) синтез ДНК считают началом профазы первого деления, а именно — прелептотенной стадией профазы. Затем следует лептотенная стадия профазы (конденсация хромосом). Причем в целом профаза мейоза (включающая шесть стадий) длится очень долго — около месяца в сперматоцитах и до нескольких десятилетий (включая длительный период покоя) в ооцитах.
В митотических же клетках, наоборот, между репликацией ДНК и началом профазы различают определенный промежуток времени, а сама профаза проходит достаточно быстро (не более нескольких часов).
2.1. Общая характеристика репликации ДНК
2.1.1. Основные принципы
Репликация ДНК имеет ряд принципиальных особенностей.
1. Субстратами, из которых синтезируются новые цепи ДНК, являются дезоксинуклеозидтрифосфаты (дНТФ), а не дезоксинуклеозид-монофосфаты (дНМФ), входящие в состав ДНК. Поэтому в ходе включения в цепь ДНК от каждого нуклеотида отщепляются 2 фосфатных остатка (в виде пирофосфата, который вскоре гидролизуется до фосфатов):

Использование именно дНТФ, а не дНМФ, объясняется энергетическими причинами: образование межнуклеотидной связи требует энергии; источником ее и служит разрыв межфосфатной связи.
2. Репликация ДНК — матричный процесс: каждая синтезируемая (дочерняя) цепь ДНК строится, используя в качестве матрицы одну из цепей исходной (родительской) ДНК.
Основой при этом является принцип комплементарности: из четырех возможных нуклеотидов (дАТФ, дГТФ, дЦТФ, дТТФ) в состав растущей цепи включается в данный момент тот, который комплементарен нуклеотиду в соответствующем положении родительской цепи.
3. Процесс (в отличие, например, от синтеза РНК) является симметричным: матрицами служат обе цепи родительской ДНК.
4. Процесс полуконсервативный: по завершении процесса исходные молекулы ДНК оказываются наполовину обновленными. В каждой из дочерних молекул одна цепь — родительская, а вторая — новосинтезированная.
5. Удлинение цепи ДНК (или отдельного ее фрагмента) всегда происходит в направлении от 5'-конца к 3'-концу. Это означает, что очередной новый нуклеотид присоединяется к 3'-концу растущей цепи.
Поскольку в любой молекуле ДНК комплементарные цепи антипараллельны, то и растущая цепь антипараллельна матричной цепи. Следовательно, последняя считывается в направлении 3' ®5¢
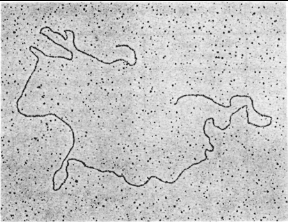
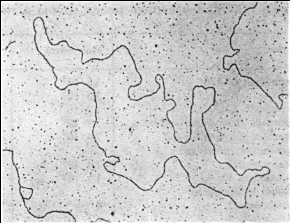
Основные механизмы репликациию. ДНК может находиться в линейной или кольцевой форме. Для эукариот и большинства прокариот характерна линейная двухцепочечная форма молекулы ДНК. Кольцевой двухцепочечной молекулой ДНК представлены бактериальные плазмиды, хромосомы некоторых бактерий, большинство митохондриальных и хлоропластных ДНК, геномы вирусов млекопитающих. Хромосома бактериофага λ на разных стадиях жизненного цикла существует то как линейная молекула, то как замкнутая кольцевая структура, то как кольцо с разрывами (рис.2.2).
 |
 |
А Б
Рис. 2.2. Электронные микрофотографии линейной (А) и кольцевой (Б) двухцепочечной ДНК фага λ.
По-видимому, никакого верхнего предела для размера кольцевой двухцепочечной молекулы ДНК не существует.
Вместе с тем не все ДНК являются двухцепочечными. Геномы некоторых мелких вирусов бактерий, растений и животных представляют собой ковалентно замкнутые кольца, состоящие только из одной цепи. Все известные одноцепочечные кольцевые ДНК относительно малы: ДНК бактериофагов φХ174 и М13 содержат примерно 5300 и 6000 нуклеотидов соответственно и имеют длину 1,5–2 мкм; длина молекул ДНК парвовирусов животных и некоторых вирусов растений составляет 2/3 и 1/2 указанных величин соответственно. Однако для репликации любой из этих вирусных ДНК совершенно необходимо превращение одноцепочечного кольца в соответствующее двухцепочечное, из которого затем образуются одноцепочечные кольцевые ДНК вирусного потомства. Более того, экспрессия генетической информации в таких геномах всегда осуществляется в фазе двухцепочечной ДНК, поскольку именно она является субстратом для транскрипции последовательности ДНК в РНК.
Для различных видов молекулы ДНК характерны отличия в механизме репликации. Выделяют двунаправленную и однонаправленную репликацию (рис.2.3).
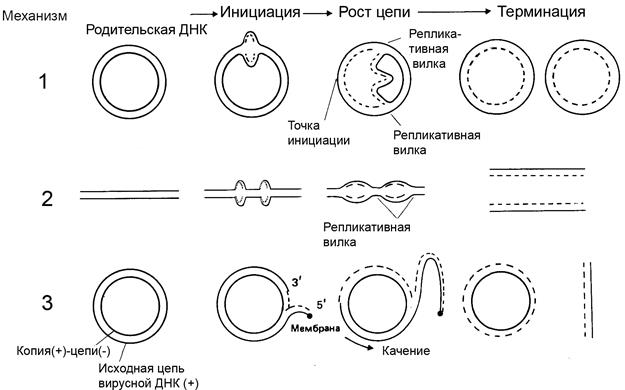
1 - Двунаправленная репликация с Q- петлей (напр. в E.coli)
2 - Двунаправленная репликация с множественной инициацией (эукариоты)
3 - Однонаправленная репликация: катящееся кольцо (напр., у фага Ф Х174).
Рис. 2.3. Механизмы двунаправленной и однонаправленной репликации
Двунаправленная репликация – наиболее распространенный механизм репликации ДНК. После инициации репликация идет одновременно в обоих направлениях вдоль цепи ДНК. Для осуществления синтеза двойная спираль родительской ДНК должна раскрутиться и ее цепи должны разойтись. В этих процессах участвуют различные белки. В линейной ДНК раскручивание осуществляется путем вращения одной цепи вокруг другой. В двухцепочечной кольцевой ДНК раскручивание и репликация ведут к образованию структуры, напоминающей кольцо с внутренней петлей. Ее называют тэта-петля, поскольку по форме она похожа на греческую букву Q. Впервые такой механизм репликации был выявлен с помощью радиоавтографии Кэрнс для процесса репликации ДНК бактерий. Для визуализации ДНК культуру E.coli выращивали на среде, содержащей радиоактивные нуклеотиды, которые включались в ДНК. Затем реплицирующуюся ДНК наносили на фотопленку, на которой и получалось изображение Q-петли.
Однонаправленная репликация – более редкое явление. Она наблюдается у некоторых вирусов, содержащих одноцепочечную ДНК. При этом репликация происходит после образования двухцепочечной формы ДНК в клетке-хозяине. Механизм, с помощью которого это осуществляется, называется механизмом катящегося кольца. При этом в одной из цепей ДНК образуется разрыв (наружная цепи на рис.2.3), и синтез новой цепи начинается с 3¢-конца этой разорванной родительской цепи с использованием второй (внутренней на рисунке) в качестве матрицы. Это приводит к вытеснению 5¢-конца наружной цепи, которая впоследствии служит матрицей для синтеза новой цепи.
Особенности механизма репликации. В настоящее время процесс репликации у прокариот достаточно изучен, в то время как многие аспекты эукариотической репликации остаются неясными. Однако с большой долей вероятности можно утверждать, что в большинстве клеток этот процесс протекает в основном одинаково.
Репликация ДНК прокариот. Ферментами, непосредственно, осуществляющими синтез новой цепи молекулы ДНК являются ДНК-полимеразы. У прокариот выделены три таких фермента: ДНК-полимераза I, II и III (pol I, pol II и pol III). Основная функция pol III- синтез новой цепи ДНК, pol I обеспечивает не только синтез, но и исправление ошибочно вставленных нуклеотидов. Полимераза II осуществляет особые, специализированные функции. Помимо 5¢®3¢-полимеразной активности все три фермента проявляют способность деградировать (разрушать) ДНК, отщепляя нуклеотиды в направлении 3¢®5¢, т.е. являются 3¢®5¢-экзонуклеазами. Pol I и pol III обладают также 5¢®3¢-экзонуклеазной активностью.
Поскольку синтез ДНК происходит на одноцепочечной матрице. Ему должно предшествовать обязательное разделение двух цепей ДНК. Участок начала расхождения цепей называется репликационной вилкой из-за характерной Y-образной формы (рис.2.4).
Именно в этой репликационной вилке ДНК-полимеразы синтезируют дочерние молекулы ДНК. Участок ДНК, в котором репликация уже завершилась, выглядит как пузырек или «глазок» в нереплицированной ДНК. Репликационные глазки образуются в тех местах, где находятся специфические последовательности – точки начала репликации (origin of replication). Они состоят примерно из 300 нуклеотидов. С ориджинами связываются инициаторные (узнающие) белки репликации. В случае бактерий такие белки называются DnaA. Инициаторные белки, обеспечив связывание ДНК-реплицирующиго комплекса ферментов, видимо, далее не перемещаются вместе с ним по ДНК.
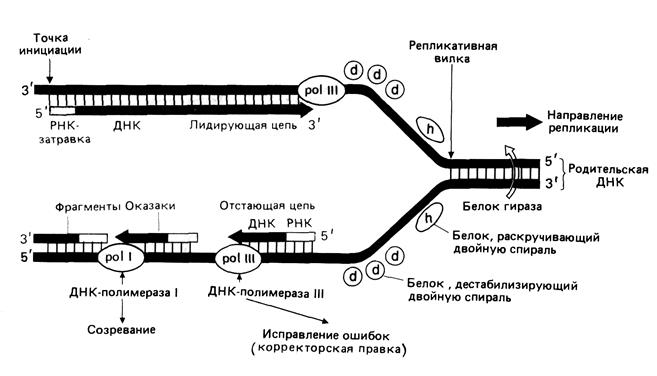
Рис.2.4. Репликация у E.coli
Для того, чтобы цепи ДНК разъединились, функционирует особый фермент – ДНК-геликаза (для бактерий это Rep-белок), который связывается с инициаторными белками. Этот фермент движется по одноцепочечной цепи ДНК и, встречая участок двойной спирали, он разрывает водородные связи между основаниями, разделяет цепи и продвигает репликационную вилку. На это затрачивается энергия гидролиза АТФ – по 2 молекулы АТФ на разделение 1 пары нуклеотидов. Одновременно происходит также вытеснение данного участка ДНК из связи с гистонами и другими хромосомными белками.
С одиночными цепями ДНК связывается специальные SSB-белки, дестабилизирующие спираль (SSB – single strand binding proteins). Они не позволяют им сомкнуться. Для того, чтобы репликационная вилка могла продвигаться вперед, вся еще не удвоенная часть ДНК должна была бы очень быстро вращаться. Белки ДНК-топоизомеразы вносят одноцепочечные или двуцепочечные разрыва, позволяющие цепям ДНК разделиться, а затем заделывают эти разрывы. Известны 2 таких фермента – топоизомераза I и топоизомераза II. Топоизомераза I разрывает одну из цепей ДНК, перенося ее проксимальный конец на себя. Это позволяет дистальному участку ДНК (от места расплетения до места разрыва) вращаться вокруг соответствующей связи целой цепи, что и предупреждает суперспирализацию нерасплетенных участков. Впоследствии концы разорванной цепи вновь замыкаются: один из них переносится с фермента на второй конец. Топоизомераза II (бактериальная топоизомераза II называется гиразой) разрывает сразу обе цепи ДНК, перенося соответствующие концы на себя. Это еще более эффективно позволяет решать проблему суперспирализации при расплетении ДНК.
Субстратом для ДНК-полимеразы являются дезоксирибонуклеозид-трифосфаты (дНТФ), полимеризующиеся на одноцепочечной матрице. ДНК-полимеразы последовательно наращивают одну нить ДНК, шаг за шагом присоединяя к ней следующие звенья в направлении от 5¢к 3¢ концу, причем выбор очередного дНТФ диктуется матрицей.
ДНК-полимеразы не могут начинать синтез ДНК на матрице, а способны только добавлять новые дезоксирибонуклеотидные звенья к 3¢-ОН- концу уже имеющейся полинуклеотидной цепи. Такую заранее образованную цепь, к которой добавляются нуклеотиды, называет затравкой (или праймером). Короткую РНК-затравку (длиной 10-60 нуклеотидов) синтезирует из рибонуклеозидтрифосфатов фермент, не обладающий корректирующей активностью и называемый ДНК-праймазой. Праймазная активность может принадлежать либо отдельному ферменту, либо одной из субъединиц ДНК-полимеразы.
Учитывая, что дочерние цепи ДНК растут только в направлении 5¢→3¢, синтез ДНК происходит непрерывно только на одной из матричных цепей. На второй (антипараллельной) цепи ДНК синтезируется сравнительно короткими фрагментами – от 100 до 1000 нуклеотидов, называемыми «фрагментами Оказаки» по имени открывшего их ученого – Тунеко Оказаки. Соответственно одна из цепей синтезируется быстрее и называется лидирующей, а вторая носит название запаздывающей.
Механизм работы ДНК-полимеразы на отстающей цепи до конца не изучен. В соответствии с одной из гипотез, полагают, что полимераза накладывает цепь ДНК, которая служит ей матрицей, саму на себя, что и обеспечивает разворот ДНК-полимеразы отстающей цепи на 180 градусов. (рис.2.5).
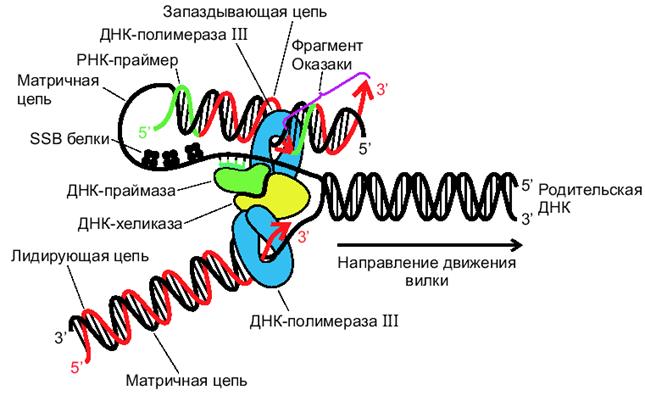
Рис. 2.5. Репликация ДНК («модель тромбона», Russell, 1998)
Такая модель репликации получила название «модель тромбона».
Генетический материал живых организмов имеет огромные размеры и реплицируется с высокой точностью. В среднем, в процессе воспроизведения генома млекопитающего, состоящего из ДНК длиной 3 млрд. пар нуклеотидов, возникает не более трех ошибок. При этом ДНК синтезируется чрезвычайно быстро (скорость ее полимеризации колеблется в пределах от 500 нуклеотидов в секунду у бактерий до 50 нуклеотидов в секунду у млекопитающих)
Высокая точность репликации, наряду с ее высокой скоростью, обеспечивается наличием специальных механизмов, осуществляющих коррекцию, т.е. устраняющих ошибки.
Суть механизма коррекции заключается в том, что ДНК-полимеразы дважды проверяют соответствие каждого нуклеотида матрице: один раз перед включением его в состав растущей цепи, второй раз перед тем, как включить следующий нуклеотид. Очередная фосфодиэфирая связь синтезируется лишь в том случае, если последний нуклеотид растущей цепи ДНК образовал правильную Уотсон-криковскую пару с соответствующим нуклеотидом матрицы.
В результате действия всех предыдущих ферментов каждая новосинтезированная цепь оказывается состоящей из фрагментов, вплотную примыкающих друг к другу. Удаление РНК-затравки с 5¢-концов всех синтезированных фрагментов осуществляет ДНК-полимераза I, действуя как экзонуклеаза. Этот же фермент на месте удаленной РНК встраивает дезоксинуклеотиды, используя при этом свою 5¢®3¢- полимеразную активность. «Сшивание» соседних фрагментов осуществляется ферментом ДНК-лигазой. Как и ДНК-полимеразы, этот фермент образует межнуклеотидную (фосфодиэфирную) связь. Но если в полимеразной реакции одним из участников является свободный дНТФ, то в ДНК-лигазной реакции оба участника - концевые дНМФ в составе «сшиваемых» фрагментов. По этой причине энергетика реакции иная, и требуется сопряженный гидролиз молекулы АТФ. Кроме того, ДНК-лигаза «сшивает» только такие одноцепочечные фрагменты, которые находятся в составе двухцепочечной ДНК.
Таким образом, процесс репликации хромосомы бактерий начинается в точке начала репликации и продолжается до тех пор, пока не удвоится вся ДНК хромосомы.
2.2.2. Репликация ДНК эукариот. Хотя молекулярно-биологические процессы, происходящие во время репликации ДНК, в основном похожи у эукариот и прокариот, между ними имеется ряд существенных различий.
1. Если бактериальная хромосома представляет собой единицу репликации – репликон, то репликация ДНК эукариотической хромосомы осуществляется посредством разделения ее на множество отдельных репликонов. Полагают, что у эукариот гомологами ориджинов начала репликации являются автономно реплицирующиеся последовательности или ARS (autonomously replicating sequences). Сначала у дрожжей Saccharomyces cerevisiae были выделены особые последовательности, которые, будучи включенными в экстрахромосомальную ДНК, обеспечивали репродукцию этих ДНК в дрожжевой клетке. Позднее такие последовательности были выделены у многих других организмов.
2. У эукариот известно 5 разных ДНК-полимераз (a- b- e- d- и g-полимераза). Из них b- и e-полимеразы участвуют в репарации ДНК, g-полимераза — в репликации митохондриальной ДНК, а a- и d-полимеразы — в репликации ядерной ДНК. При этом альфа-полимераза контролирует рост лидирующей цепи, а дельта полимераза – запаздывающей.
По некоторым предположениям, a--полимераза связана и с праймазой, и с d-полимеразой, а последняя, в свою очередь, — с белком PCNA (от англ. Proliferation Cell Nuclear Antigen).
Данный белок выполняет роль «прищепки», которая крепит комплекс полимераз к реплицируемой цепи ДНК. Считается, что в «застегнутом» состоянии он, как кольцо, обхватывает цепь ДНК. Тем самым предупреждается преждевременная диссоциация полимераз от данной цепи.
3. Созревание фрагментов Оказаки у эукариот требует удаления РНК-затравок с помощью 5'®3'-экзонуклеазы ( белковые факторы FEN-1 или MF-1) и РНКазы Н1, а также ковалентного соединения фрагментов друг с другом под действием ДНК-лигазы I .
По эукариотической хромосоме в каждый момент времени может двигаться независимо друг от друга множество репликационных вилок. Остановка продвижения вилки происходит только при столкновении с другой вилкой, движущейся во встречном направлении, или по достижении конца хромосомы. В результате вся ДНК хромосомы в короткий срок оказывается реплицированной.
Насколько велики репликоны и как много их в геноме? Трудность определения размеров и числа репликонов заключается в том, что трудно выделить индивидуальные «глазки» репликации. Всегда остается возможность, что наблюдаемый глазок является результатом слияния двух соседних репликонов. Чтобы обойти это препятствие, подбирают стадию репликации, когда число «глазков» максимально и они еще не начали сливаться. Затем в участке ДНК, содержащем несколько глазков, измеряют расстояние между начальными точками начала репликации (т.е. между средними точками смежных репликонов). Скорость движения вилки репликации определяют по максимальной длине радиоактивно меченого следа реплицирующейся ДНК в единицу времени.
Дата добавления: 2016-02-09; просмотров: 4483;