История исследования ДНК. Биологические свойства ДНК. Химический состав молекулы ДНК
Ко времени открытия структуры ДНК история изучения нуклеиновых кислот насчитывала уже около восьмидесяти лет. Честь их открытия принадлежит швейцарскому биохимику Фридриху Мишеру, который в 1868—1872 гг. выделил из ядер клеток гноя и спермы лосося новое фосфорсодержащее вещество, названное им нуклеином (от греч.— ядро). Примерно в те же годы Мендель старался убедить ученый мир в значении своей работы. До середины XX столетия никто не предполагал, что эти два открытия столь тесно связаны между собой. Работа Менделя пребывала в забвении до 1901 г., а результаты исследований Мишера в подробном изложении были опубликованы после его смерти — в 1890 г. Незадолго до этого, в 1889 г., немецкий химик Р. Альтман впервые получил свободный от белков нуклеин Мишера и предложил назвать его нуклеиновой кислотой.
К этому времени А. Коссель выделил основные составные части нуклеина: содержащие азот вещества — аденин и гуанин, фосфорную кислоту и соединения из группы углеводов. Впоследствии удалось установить, что в природе существует два типа нуклеиновых кислот ДНК и РНК (у последней сахар— дезоксирибоза — заменен на рибозу, а одно из четырех азотистых оснований — тимин — на урацил). Так как ДНК выделяли в основном из тимуса теленка, а РНК — из дрожжей и растений, то долгое время бытовало представление о том, что ядра клеток животных содержат только ДНК, а ядра клеток растений — только РНК. Только к середине 30-х годов было доказано, что ДНК и РНК содержатся в каждой живой клетке. С развитием методов цитохимии и гистохимии, а также методов фракционирования субклеточных структур к концу 40-х годов было установлено, что ДНК локализуется преимущественно в ядре, а РНК в цитоплазме клеток.
К началу 50-х годов были установлены основные принципы химического строения нуклеиновых кислот. Была выяснена структура их мономеров — нуклеозидов и нуклеотидов, и доказано, что и в ДНК, и в РНК нуклеотидные остатки связаны только 3' — 5'-фосфодиэфирной связью.
До 40-х годов исследование нуклеиновых кислот считалось весьма скучным и вообще бесперспективным занятием. Так продолжалось до 1944 г., когда Эйвери, Мак-Леод и Мак-Карти установили, что дезоксирибонуклеиновая кислота является носителем генетической информации. Это выдающееся открытие, приведшее к установлению химической природы генов. Его история ведет свое начало с 1928 г., когда Фред Гриффит провел эксперименты с заражением мышей пневмококками. Пневмококки вызывают пневмонию у человека и других чувствительных к ним млекопитающих. Они обычно окружены слизистой блестящей оболочкой — полисахаридной капсулой. Этот наружный слой имеет существенное значение для проявления патогенности бактерий. Мутанты, лишенные полисахаридной оболочки, не патогенны. Патогенные бактерии дикого типа обозначают буквой S (от англ. smooth — гладкий), так как они образуют гладкие колонии, а мутантные бактерии, не имеющие капсулы, — буквой R (от англ. rought — шероховатый), так как они образуют шероховатые колонии.
Гриффит обнаружил, что непатогенный мутант можно трансформировать в патогенную S-форму следующим образом. Он инъецировал мышам смесь живых бактерий R-формы и убитых нагреванием пневмококков S. При этом были получены поразительные результаты. Оказалось, что указанная смесь вызывала гибель мышей, хотя ни живые пневмококки, ни убитые нагреванием пневмококки, инъецированные порознь, смерти мышей не вызывали. В крови погибших мышей содержались живые S пневмококки. Следовательно, убитые нагреванием пневмококки каким-то образом трансформировали живые R- и живые S-пневмококки. Это изменение стабильно наследовалось: трансформированные пневмококки давали патогенное потомство S-формы.
Впоследствии R-S-трансформацию удалось воспроизвести в бесклеточной системе (in vitro). Некоторые R-клетки в растущей культуре трансформировались в S-форму при добавлении бесклеточного экстракта убитых нагреванием пневмококков. Это открытие позволило установить химическую природу трансформирующего фактора.
Эйвери с сотр. в своих классических исследованиях показали, что трансформирующий фактор - это дезоксирибонуклеиновая кислота. Новое доказательство трансформирующей активности ДНК было получено несколько позже, когда удалось очистить фермент дезоксирибонуклеазу, разрушающую ДНК. Было показано, что добавление этого фермента необратимо инактивирует трансформирующий фактор.
Публикация выводов Эйвери с сотр. в 1944 г. была встречена с большим удивлением и недоверием, так как едва ли кто-либо ранее придавал ДНК информационную роль. Вездесущему присутствию ДНК в хромосомах большей частью приписывали чисто физиологическую или структурную роль. В то же время считали, что именно хромосомный белок придает генам информационную роль, поскольку еще в начале XX в. были выявлены различия в специфичности белков у различных организмов. Авторы понимали трудность обоснования генетической роли ДНК и в заключительной части своей работы высказали следующее утверждение: «Если результаты представленного исследования о природе трансформирующего начала подтвердятся, то придется признать, что нуклеиновые кислоты обладают биологической специфичностью, химическая основа которой еще не установлена».
В 1952 г. Альфред Херши и Марта Чейз доказали генетическую роль ДНК в совершенно иной системе — при изучении вируса (бактериофага), заражающего бактерию Е. соli. Когда фаги добавляют к бактериальной культуре, они адсорбируются на наружной поверхности бактерии и вводят в нее определенное вещество, в результате чего примерно через 30 мин бактерия разрывается (лизирует), высвобождая большое число новых фаговых частиц — потомков, адсорбированных фагов.
Эксперимент был поставлен следующим образом. Бактерии инфицировали фагом Т2, у которого радиоактивным изотопом метили либо ДНК-компоненты (изотопом фосфора — 32Р), либо белковые компоненты (изотопом серы — 35S). Фаги смешивали с бактериями и неадсорбированные частицы удаляли центрифугированием. Затем инфицированные бактерии энергично встряхивали и разделяли полученный препарат на две фракции путем центрифугирования. Одна фракция содержала пустые фаговые оболочки, отделившиеся от клеточной стенки бактерий, другая — сами бактерии. Анализ фракций показал, что 35 метка была связана с оболочками фага. Большая же часть метки 32Р оказалась внутри инфицированных бактерий. В потомстве фага после инфицирования было найдено примерно 30% исходной метки 32Р. А от исходного белка в фаговом потомстве обнаружили лишь менее 1%. Этот эксперимент прямо показывает, что родительская фаговая ДНК проникает в бактерию и затем становится частью фагового потомства. Именно так должно происходить наследование генетического материала.
Эксперименты Херши и Чейз убедительно подтвердили факты, открытые восемью годами раньше Эйвери с сотр. на другой системе, и результаты работы были сразу восприняты как доказательство генетической роли ДНК.
До 1950 г. доминировала тетрануклеотидная теория структуры ДНК, рассматривающая полинуклеотид как монотонную макромолекулу, состоящую из многократно повторяющихся наборов четырех нуклеотидов. Такая молекула, очевидно, не способна выполнять роль носителя генетической информации. В 1948 г. Хочкисс и Эрвин Чаргафф применили тогда еще новый метод хроматографии на бумаге для разделения и количественной оценки компонентов нуклеиновой кислоты. Этот метод давал возможность проводить более точный анализ нуклеотидных оснований, высвобождающихся при полном гидролизе ДНК. Анализы показали, что в противоположность требованиям тетрануклеотидной теории четыре азотистых основания не обязательно присутствуют в ДНК в точно равных соотношениях. Чаргафф проанализировал образцы ДНК, выделенной из разных организмов, и показал, что молярные соотношения оснований в молекуле ДНК могут варьировать в широких пределах в зависимости от ее биологического источника. Но если она не монотонна, то тогда вполне возможно, что именно состав ее оснований и обусловливает ее биологическую специфичность.
К 1952 г. удалось сформулировать теорию, объясняющую, каким образом ДНК может осуществлять перенос генетической информации в опытах с трансформацией. Основное положение этой теории сводилось к следующему: если молекула ДНК содержит генетическую информацию, то последняя определяется не чем иным, как специфической нуклеотидной последовательностью четырех нуклеотидных оснований в полинуклеотидной цепи.
В 1953 г. Джеймс Уотсон и Френсис Крик установили трехмерную (вторичную) структуру ДНК и сразу же предложили механизм ее репликации (удвоения). Это блестящее достижение стоит в ряду важнейших событий в истории биологии, так как оно открыло путь к пониманию функции гена на молекулярном уровне. Не случайно 1953 год считают годом рождения молекулярной биологии.
Две группы фактов легли в основу модели ДНК. Первая связана с успешным использованием рентгеноструктурного анализа для изучения биологических макромолекул. Одним из первых исследователей, высказавших соображения о трехмерной структуре ДНК, был Астбюри (именно он ввел в 1940 г. термин «молекулярная биология»), который предположил, что полимер ДНК представляет собой стопку уложенных один над другим нуклеотидов. Его измерения показали также, что нуклеотидные остатки, ориентированные перпендикулярно длинной оси молекулы, располагаются вдоль оси через каждые 3,4 Å (1 ангстрем = 10-10 м). Группа исследователей, продолжившая рентгеноструктурные исследования ДНК, начатые Астбюри, и работавшая под руководством Уилкинса, достигла важного методического решения: им удалось приготовить высокоориентированные нити ДНК, на основе которых получили рентгенограмму, показывающую множество ранее не проявлявшихся деталей.
Уотсон и Крик к тому времени рассмотрели несколько возможных вариантов структуры ДНК, однако из-за плохого качества рентгенограмм им не удалось прийти к каким-либо определенным выводам. Рентгенограмма, полученная Розалиной Франклин, сотрудницей Уилкинса, помогла узнать недостающие детали, и в течение нескольких недель вопрос о структуре ДНК был решен.
Определяющую роль в установлении структуры ДНК сыграли факты, полученные Чаргаффом при анализе нуклеотидного состава ДНК различных организмов. В его докладе, сделанном в 1950 г., можно найти следующее утверждение: «Полученные результаты служат опровержением тетрануклеотидной гипотезы. Следует, однако, отметить — хотя трудно еще сказать, не является ли это чистой случайностью,— что во всех изученных до сих пор дезоксирибонуклеиновых кислотах молярные отношения пуринов к пиримидинам в целом, а также аденина к тимину и гуанина к цитозину близки к 1. В этом утверждении впервые была сформулирована важная структурная особенность ДНК, подтвержденная последующими анализами: несмотря на довольно широкое разнообразие в составе (у бактерий молярная доза Г + Ц варьирует от 26 до 74%), проявляемое различными типами ДНК, молярное содержание тимина равно молярному содержанию аденина, так же как содержание гуанина равно содержанию цитозина.
В апреле 1953 г. Уотсон и Крик опубликовали небольшую статью, в которой постулировали структуры ДНК. В том же выпуске Уилкинс с сотр. опубликовали данные рентгеноструктурного анализа, подтверждающие правильность предложенной модели.
Биологические свойства ДНК. ДНК – полимерная макромолекула, состоящая из тысяч мономеров и имеющая большую молекулярную массу (6х106-6х1012 Да). Мономерами являются нуклеотиды. Участки дезоксирибонуклеиновой кислоты имеют специфическую последовательность нуклеотидов, которая кодирует последовательность аминокислот в полипептидах и рибонуклеотидов в РНК.
Молекула ДНК имет ряд уникальных свойств:
1. Она способна удваиваться во время S-периода интерфазы. Этот процесс называется репликацией. В процессе митоза копии ДНК равномерно распределяются между дочерними клетками.
2. ДНК служит матрицей для образования своих информационных копий в виде РНК. Этот процесс называется транскрипцией. Копия ДНК в виде РНК непосредственно принимает участие в синтезе белков.
3. В случае повреждения ДНК способна к восстановлению структуры, благодаря чему поддерживается консервативность генетической информации. Этот процесс называется репарацией.
Все три вышеназванные процессы происходят с участием ферментов.
4. ДНК способна к мутации – изменению последовательности нуклеотидов.
Химический состав молекулы ДНК. В состав мономерной единицы ДНК- нуклеотида входит :
· азотистое основание
· моносахарид
· один фосфатный остаток

Азотистые основания представляют собой производные одного из двух соединений – пурина или пиримидина. Пуриновыми производными в ДНК являются аденин (А) и гуанин (G), а пиримидиновыми – цитозин (С) и тимин (Т). Азотистое основание присоединяется к моносахариду с помощью b-N-гликозидной связи, соединяющей С1′-атом моносахарида с N1-атомом пиримидинового производного или N9-атомом пуринового (рис.1.1).
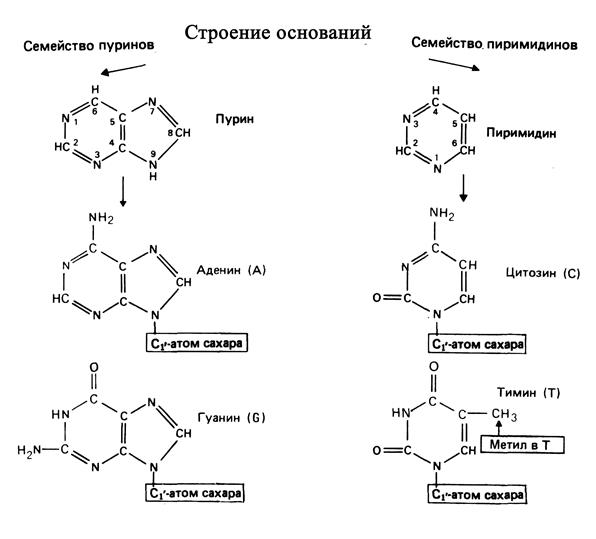
Рис. 1.1. Строение азотистых оснований молекулы ДНК
Моносахарид, входящий в состав нуклеотида – это пентоза в форме b-D-2-дезоксирибозы (рис.1.2).
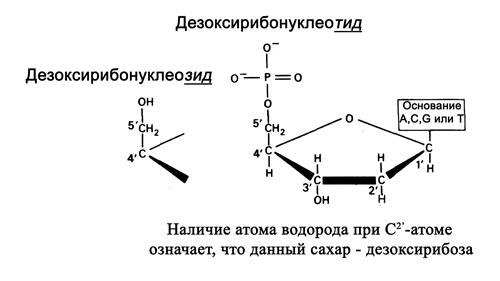
Рис.1.2. Строение сахара в ДНК
В дезоксирибозе в отличии от рибозы гидроксильная (-ОН) группа при 2′-углеродном атоме пентозы замещена на атом водорода.
Азотистое основание, связанное с пентозой, называется нуклеозидом: для ДНК это,соответственно, дезоксиаденозин, дезоксигуанозин, дезоксицитидин и дезокситимидин.
Исходя из этого, составляются и названия нуклеотидов ДНК:
дАМФ-дезоксиаденозинмонофосфат
дГМФ- дезоксигуанозинмонофосфат
дЦМФ- дезоксицитидинмонофосфат
дТМФ – дезокситимидинмонофосфат
Структура ДНК
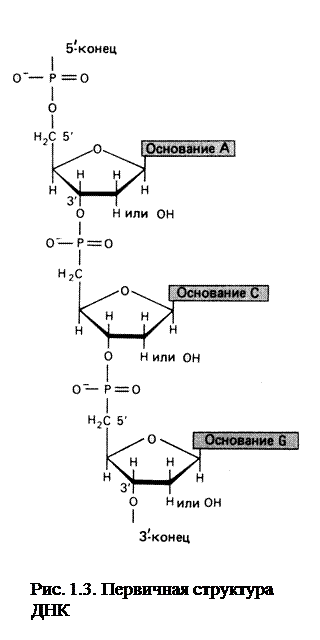
Подобно белкам, ДНК имеет первичную, вторичную и третичную структуры.
Каждая из полинуклеотидных цепей ДНК построена из ковалентно связанных дезоксирибонуклеотидных звеньев (рис.1.3). Причем отдельные нуклеодиты в цепи соединяются между собой с помощью фосфодиэфирных мостиков, связывающих 5′ -гидроксильную группу пентозы одного нуклеотида и 3′-гидроксильную группу пентозы следующего. В результате образуется регулярная основная цепь фосфат-сахар-фосфат-сахар – и т.д. (сахарофосфатный остов). При этом азотисные основания как бы «выступают» из сахарофосфатного остова.
Порядок следования оснований вдоль цепи называется первичной структурой ДНК.
Полинуклеотидная цепь обладает полярностью. Концевой нуклеотид на одном конце цепи имеет свободную (не участвующую в образовании межнуклеотидной связи) 5′-группу, на другом конце 3′-группу. Согласно принятому соглашению, последовательность нуклеотидов читается в направлении от 5′ - к 3′- углеродному атому пентозы, т.е. от 5′ - к 3′-концу цепи.
Вторичная структура ДНК представляет собой свернутые в спираль две комплементарно взаимодействующие и антипараллельные полинуклеотидные цепи (рис.1.4). Образование вторичной структуры ДНК возможно вследствие проявления эффектов комплементарности и стекинг-взаимодействий.
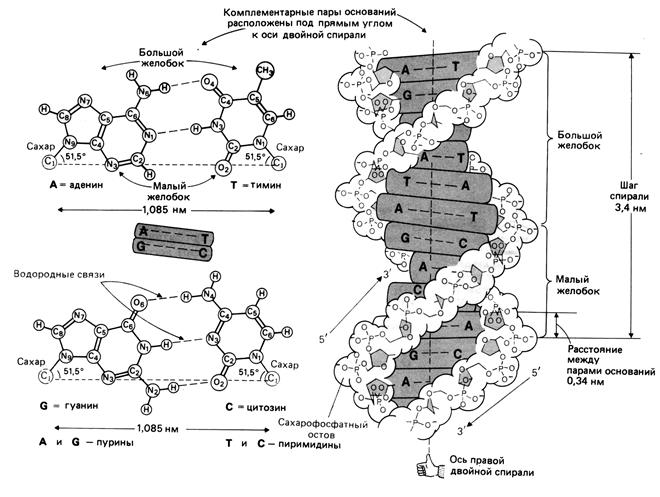
Рис. 1.4. Вторичная структура ДНК
Согласно трехмерной модели, предложенной Уотсоном и Криком, сахаро-фосфатный остов располагается по периферии двойной спирали, а азотистые основания находятся внутри, и их плоскости перпендикулярны оси спирали. Между основаниями образуются специфические водородные связи, в результате чего осуществляется так называемое уотсон-криковское спаривание. Аденин всегда образует две водородные связи с тимином, а гуанин образует три водородные связи с цитозином. Таким образом, более объемные пурины всегда спариваются с пиримидинами, имеющими меньшие размеры. Это приводит к тому, что расстояния между С1'-атомами дезоксирибозы в двух цепях оказываются одинаковыми для АТ- и GC-пар и равными 1,085 нм. В результате АТ- и GС-пары включаются в двойную спираль без какого-либо заметного изменения геометрии остова.
Диаметр двойной спирали равен примерно 2,0 нм. В двойной спирали ДНК образуются желобки. Это связано с тем, что сахарофосфатный остов расположен дальше от оси спирали, чем основания. В двойной спирали имеются два желобка-большой и малый.
Стабильность двойной спирали обусловлена разными взаимодействиями. Отчасти за нее ответственны водородные связи между основаниями. Однако, по-видимому, более важную роль играет межплоскостное взаимодействие-стэкинг. При этом обеспечиваются не только выгодные вандер-ваальсовы контакты между атомами, но и возникает дополнительная стабилизация благодаря перекрыванию p-орбиталей атомов контактирующих оснований. Стабилизация осуществляется также за счет благоприятного гидрофобного эффекта, проявляющегося в том, что неполярные основания защищены от непосредственного контакта с растворителем. Напротив, сахарофосфатный остов с его полярными группами и заряженными атомами экспонирован, что также стабилизирует структуру.
Для вторичной структуры характерно явление полиморфизма ДНК. Изменение физиологических условий приводит к переходу одной формы (конформации) ДНК в другую, что позволяет дополнительно регулировать работу генов. Т.о. полиморфизм ДНК-это способность двойной спирали принимать различные конформации. Рентгеноструктурные исследования кристаллов полинуклеотидов выявили четыре основных типа структур – А-, В-С- и Z-формы ДНК (рис.1.5).
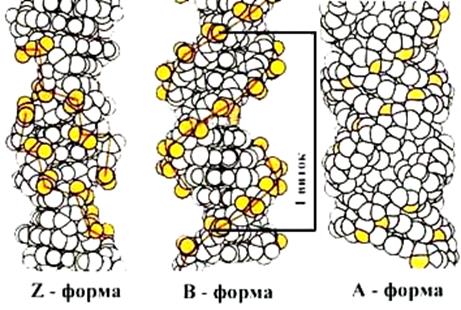
Рис. 1.5. Различные конформации ДНК
В-ДНК- этостандартная уотсон-криковская структура, в которой плоскости пар оснований перпендикулярны оси двойной спирали.
Соседние комплементарные пары повернуты друг относительно друга приблизительно на 360. В соответствии с последними данными на один виток спирали приходится 10,5 пар оснований, и шаг спирали соответственно равен 36 Å. Это правозакрученная спираль.
А-ДНК образуется при высушивании волокон В-ДНК, в водных растворах с повышенным содержанием калия и натрия или при добавлении спирта. Расстояние между нуклеотидами вдоль оси спирали составляет 2,56 Å. Высота витка спирали 28Å. На виток спирали здесь приходятся 11 пар оснований. Пары оснований в A-ДНК, так же как и в B-ДНК, почти плоские, но в этой форме они наклонены на 20 град. относительно перпендикуляра к оси спирали и смещены относительно оси на 4,7Å, так что ось попадает в большой желобок. Это приводит к появлению полости в центре структуры диаметром 5Å (рис. 1.6)
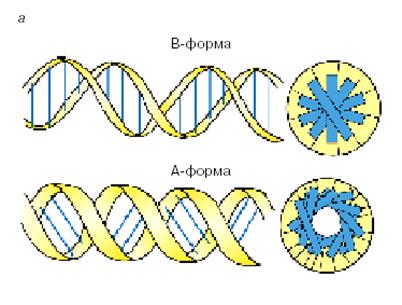
Рис. 1.6. А- и В- формы ДНК
С-ДНК также является правозакрученной спиралью. В этой форме ДНК шаг спирали составляет 31Å. На один виток спирали приходится 9,3 пар оснований. Угол наклона плоскости пар оснований к нормали оси спирали - 60.
Z- ДНК - левоспиральная (в отличие от A-, B- и С-ДНК ) конформация ДНК. Она была открыта в 1979 г. при исследовании структуры гексануклеотида d(CG)3. Если полинуклеотид poly(dG-dC) поместить в водный раствор с высокой концентрацией MgCl2, NaCl или спирта, то образуется левая двойная спираль Z-ДНК. Повторяющейся единицей спирали является не пара нуклеотидов, а двойка соседних пар. Высота витка в Z-форме -44,5 Å, на виток приходится 12 пар нуклеотидов. Плоскости оснований примерно перпендикулярны оси спирали. Буква Z указывает на зигзагообразную форму сахарофосфатного остова ДНК в этой форме (рис.1.5).
В клетке ДНК обычно находится в В-форме, но отдельные ее участки могут находиться в А-, С-, Z- или даже иных конформациях.
У эукариот ДНК находится в специальном клеточном компатменте – ядре. В среднем на интерфазное ядро млекопитающих приходится около 2 м ДНК, которая локализуется в сферическом ядре со средним диаметром около 10 мкм. Т.е. молекула ДНК должна быть уложена с коэффициентом упаковки 1х103-1х104 в упорядоченную структуру обеспечивающую репликацию и транскрипцию молекулы. Третичная структура ДНК в эукариотических клетках отличается тем, что многократная спирализация ДНК сопровождается образованием комплексов с белками. Такая укладка обеспечивает формирование ядерного хроматина или в максимально конденсированном виде – метафазных хромосом (рис.1.7).
При образовании третичной структуры ДНК возможно образование трех- и четырехнитевых участков. Образование трехнитевых участков происходит благодаря так называемым Хугстэновским взаимодействиям (Hoogsteen base pairs), когда одновременно взаимодействуют три основания: А-А-Т, Т-А-Т, Г-Г-Ц, Ц-Г-Ц. Аналогичным образом происходит образование тетрамерных участков ДНК. Точный биологический смысл появления трех- и четырехнитевых участков ДНК пока не выяснен. Имеются только лишь предположения о том, что такие участки возникают в местах, наиболее ответственных за процессы репликации и транскрипции.
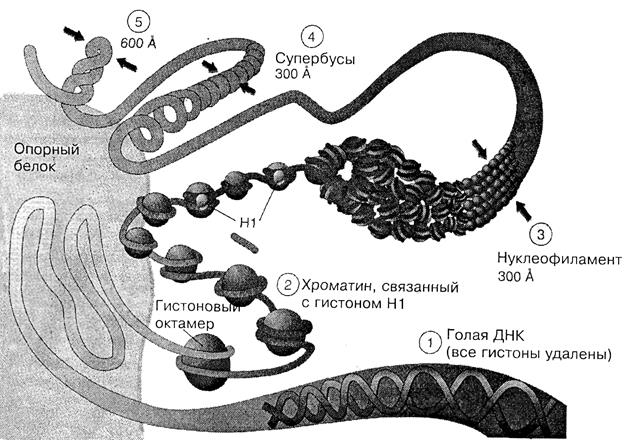
Рис. 1.7. Организация хромосомного материала
С хромосомными нитями ДНК связываются главным образом так называемые гистоны – небольшие сильно щелочные белки. Примерно 60-80% хромосомных белков представлены гистонами. Гистоны обогащены аминокислотами с основными (аргинин, лизин) и гидрофобными (валин и др.) радикалами (табл.1.1).
Таблица 1.1
Общие свойства гистонов млекопитающих
| Гистон | Молекулярная масса | Основные аминокислоты, % | Кислые аминокисло-ты, % | Отношение основных аминокислот к кислым | |
| лизин | аргинин | ||||
| H1 H2A H2B H3 H4 | 23 000 13 960 13 770 15 340 11 280 | 5,4 1,4 1,7 1,8 2,5 |
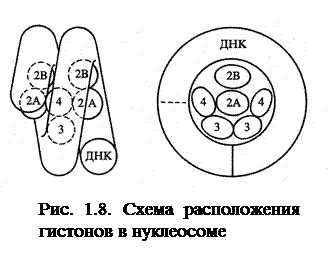
Благодаря основным радикалам, гистоны взаимодействуют с ДНК, а благодаря гиброфобным радикалам – друг с другом. Эти взаимодействия приводят к образованию нуклеосом. Основа нуклеосомы - глобула из 8 белковых молекул (октамер) она включает по 2 молекулы гистонов 4-х видов (Н2А Н2В Н3 и Н4). Вокруг одной такой глобулы молекула ДНК делает примерно 2 оборота, что и образует в итоге нуклеосому (рис.1.8).
Участки ДНК, «намотанные» на гистоновые октамеры, имеют длину в 140 нуклеотидных пар (н. п.) и называются коровыми (соrе-ДНК, или nDNA).
Нуклеосомы расположены на молекуле ДНК не вплотную: между ними имеются линкерные (соединительные) участки (iDNA) длиной 60 н. п. С каждым линкерным участком связана 1 молекула еще одного вида гистонов — Н1.
Следовательно, всего в хромосомах встречаются гистоны 5 видов, а период нуклеосомной организации составляет 200 н. п.
Молекула ДНК участвует в образовании очень большого числа нуклеосом (в среднем 600 000). В результате на данном уровне организации каждая хромосома (не считая негистоновых белков) представляет собой длинную нить «бусинок» — нуклеосом диаметром 10 нм. По сравнению с молекулой ДНК, длина нуклеосомной нити примерно в 6,2 раза меньше.
В интерфазном ядре хромосомы не различимы, а воспринимаются все вместе как хроматин. При этом выделяют гетеро- и эухроматин.
Гетерохроматин — сильно конденсированные и потому функционально неактивные участки хромосом. Они имеют вид плотных глыбок и интенсивно красятся базофильными красителями. Многие глыбки находятся на периферии ядра и прилежат к ядерной оболочке.
Напротив, эухроматин — функционально активные, практически деконденсированные и потому светлые участки хромосом, расположенные между глыбками гетерохроматина.
Нуклеосомный уровень организации имеется, видимо, и в гетеро-, и в эухроматине. Но в тех локусах эухроматина, на которых в данный момент времени функционируют ферментные комплексы (репликации, репарации или транскрипции), как полагают, ДНК высвобождается из взаимодействия с гисто-нами. Т. е. здесь нуклеосомная организация временно исчезает — с тем, чтобы впоследствии вновь восстановиться.
В отличие от этого, в гетерохроматине к нуклеосомному уровню добавляются последующие уровни укладки хромосомы. Считается, что нуклеосомная нить закручивается в спираль типа соленоида, а та, возможно, образует суперспираль. В этих процессах, видимо, ключевую роль играет гистон Н1. В итоге формируется хроматиновая нить диаметром 30 нм.
Хроматиновая нить короче нуклеосомной примерно в 18 раз и короче упакованной в ней молекулы ДНК в 6,2 х 18 @ 100 раз.
В свою очередь, хроматиновые нити образуют петли, которые собираются в розетки, где основания петель крепятся к белкам ядерного матрикса. В гетерохроматине такие группы петель более или менее плотно прилежат друг к другу.
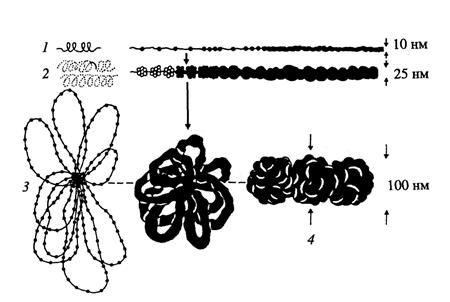
Рис. 1.9. Схема начальных уровней компактизации хроматина
1- нуклеосомный; 2- нуклеомерный (30-нанометровая фибрилла); 3- хромомерный (петлевой домен); 4 - хромонемный
Наибольшей компактизации хромосомы достигают в процессе митоза (на стадии метафазы). Точная укладка хромосомных нитей при этом неизвестна.
По одной из версий, хроматиновая нить многократно складывается по длине хромосомы. Поэтому при микроскопии на поперечном срезе обнаруживается около 100 хроматиновых нитей (представляющих собой сечения одной и той же нити). Это приводит уже к 600-кратной компактизации ДНК (рис.1.9).
Кроме того, петли хроматиновой нити имеют длину не всей хромосомы, а лишь отдельных ее сегментов — хромомеров. Это объясняет возможность сегментации хромосом при тех или иных воздействиях.
Дата добавления: 2016-02-09; просмотров: 3212;