Дифференцировка и детерминация
Раздел генетики, изучающий генетические основы индивидуального развития (онтогенеза), называется феногенетикой.Онтогенез включает увеличение массы организма (рост) и структурно-функциональную дифференциацию составляющих его клеток. Понятия «рост» и «развитие» применимы как к одноклеточным, так и к многоклеточным эукариотическим организмам, однако специфика онтогенеза тех и других обусловлена глубокими эволюционными отличиями между ними, связанными с возникновением многоклеточности. В данной теме рассматриваются в основном вопросы, касающиеся генетической регуляции онтогенеза многоклеточных животных.
Индивидуальное развитие начинается с оплодотворенной яйцеклетки. Однако уже организм новорожденного ребенка содержит около 1014 клеток, а тело взрослого человека состоит из 1015-1016 клеток. В результате процессов, происходящих в онтогенезе, формируются органы и ткани, выполняющие, как правило, ограниченное число функций. Структура клеток тканей взрослого организма отличается, и весьма заметно, от структуры яйцеклетки и приспособлена к выполнению тканеспецифических функций, возникающих в ходе дифференцировки. Дифференцировка — это процесс формирования структурно-функциональной организации клеток многоклеточных животных и растений, в результате которого клетки приобретают способность к выполнению определенных функций в сложном организме. Дифференцированное состояние в норме стабильно: клетки нервной ткани не превращаются в печеночные или эпителиальные клетки кишечника, и наоборот.
Советский биолог А. А. Заварзин открыл основную тенденцию в эволюции тканевых клеток: по мере усложнения организации их носителей они все больше и больше становятся частями целого, теряют самостоятельность и в своих проявлениях целиком зависят от надклеточных регуляционных систем: внутри- и межтканевых отношений, гуморальных и нервных факторов. Другими словами, соматические клетки животных эволюционируют как субъединицы целостного организма. Отсюда ясно, что этот принцип был бы нарушен, если бы происходило постоянное превращение одних клеток в другие. В свою очередь, это нарушило бы гомеостатические реакции организма и резко снизило бы его устойчивость к внешним факторам. Взаимодействие клеток тканей и надклеточных регуляционных систем основано на компетентности, т. е. восприимчивости клеток к регуляционным влияниям. Фактически все основные функции дифференцированных клеток контролируются организмом. Этот принцип нарушается в злокачественных опухолях.
Конечная дифференцировка часто связана с утратой способности клеток к размножению – пролиферации. Активная пролиферация и функционирование — процессы в норме, как правило, взаимоисключающиеся. Это, вероятно, и служит причиной того, что, например, нервные клетки млекопитающих делятся последний раз в эмбриональном и раннем постэмбриональном периодах, а клетки нервных ганглиев дрозофилы — в личиночной стадии. При развитии взрослого организма формируются комплексы, состоящие из многих нервных клеток, выполняющих разные функции, начиная от чувствительного нейрона и кончая двигательным. Если бы нервные клетки постоянно делились во взрослом организме, то поддержание целостности этих комплексов было бы невозможно и это, несомненно, имело бы катастрофические последствия для нервной регуляции.
Итак, дифференцированное состояние проявляется в специфическом «портрете» соматических клеток и их функциональной характеристике. Однако в пролиферирующих тканях клетки дифференцированы в разной степени. Например, в эпителии на вершине ворсинки находятся клетки, достигшие конечной стадии дифференцировки, тогда как клетки в криптах — камбиальных отделах кишечного эпителия — морфологически сильно отличаются от клеток вершины ворсинок. Однако из недифференцированных клеток крипт могут возникнуть только эпителиальные клетки кишечника. В этом случае можно утверждать, что клетки крипт детерминированы, т. е. могут развиваться только в каком-либо определенном направлении. Детерминация начинается в раннем эмбриогенезе и постепенно сужает число возможных превращений клеток до одного какого-либо дифференцированного состояния или очень немногих.
В опытах по пересадке ядер на амфибиях, проведенных в 50-е годы, было показано, что полноценное развитие может быть обеспечено только ядрами, взятыми на самых ранних стадиях развития животных. Такие ядра называются тотипотентными, т. е. способными повторить все стадии развития организма и дать все типы клеток. Ядра, взятые из сформировавшихся первичных зародышевых тканей, такую способность теряют. Например, эктодермальные ядра, пересаженные в энуклеированную яйцеклетку, приводили к развитию зародыша с дефектной энтодермой, и, напротив, энтодермальные ядра не способны образовывать эктодерму. Позднее было показано (см. ниже), что при определенных экспериментальных условиях тотипотентность ядер клеток даже дифференцированных тканей может быть восстановлена, однако важно подчеркнуть, что в норме процессы сужения потенций ядер соматических клеток развиваются необратимо.
Интересные факты, касающиеся детерминации, были получены на дрозофиле. Как известно, многие насекомые, в том числе и дрозофила, развиваются путем полного метаморфоза. Это означает, что на стадии куколки происходит лизис личиночных тканей, кроме нервных ганглиев, гонад и имагинальных дисков. Из имагинальных дисков путем пролиферации и дифференцировки развиваются органы взрослой мухи. Они состоят из дифференцированных постмитотических неделящихся клеток. Има-гинальные диски можно выделить из личинки и трансплантировать в брюшко взрослых самок. Там под влиянием гормонов реципиента они пролиферируют без дифференцировки. Если их возвратить в полость тела личинки незадолго до окукливания, то они дифференцируются в строгом соответствии со своим происхождением. Так, имагинальные диски, определяющие развитие глаз, даже если они пересажены в необычное место, например в брюшную часть тела другой личинки, развиваются в глаза.
2.Эпигеномная наследственность
Возникает вопрос: каков механизм поддержания стабильности дифференцированного состояния; почему клетки, детерминированные в определенном направлении, сколько бы раз они ни делились, сохраняют свою специфичность? Вейсман полагал, что в основе детерминации лежат неравнонаследственные деления. Носителем полной генетической информации, т.е. детерминантов (генов) всех признаков взрослого организма, является оплодотворенная яйцеклетка. Тканевые клетки получают набор генов, соответствующих их структуре и функциям. Это означает, что в нервных клетках нет генов гемоглобина, а в клетках печени — генов, кодирующих белки мышц. В качестве подтверждения своей гипотезы Вейсман использовал данные по диминуции хроматина у лошадиной аскариды. Он полагал, что диминуция – это как раз тот процесс, при котором тканевые клетки освобождаются от лишнего генетического материала. Эта гипотеза оказалась ошибочной. Диминуционные деления (их обычно одно или два) происходят в раннем эмбриогенезе (3-7 делений дробления), т. е. тогда, когда ткане-специфические гены еще не начинают функционировать. Собственно говоря, диминуция — это первый акт детерминации, разделяющий зародышевые половые и соматические клетки. Все типы тканевых клеток развиваются после диминуционных делений. Нельзя не отметить, что диминуция наблюдается у ничтожного числа из ныне известных видов животных. Данные цитогенетики показывают, что у видов, у которых диминуция отсутствует, кариотины всех тканевых клеток одинаковы; цитофотометрия свидетельствует о том, что клетки различной дифференцировки по содержанию ДНК не различаются; наконец, данные молекулярной гибридизации нуклеиновых кислот указывают на идентичность спектра нуклеотидных последовательностей в клетках разных тканей. Следовательно, гены гемоглобина присутствуют не только в эритро-идных клетках, где они активно функционируют, но и в клетках мозга, печени, почек и других тканей.
Таким образом, дифференциация происходит на основе неизменного в количественном отношении генома, сохраняющего полный спектр всех своих компонентов. Однако допускается, что в процессе дифференцировки отдельные гены избирательно повреждаются и в данных тканевых клетках уже никогда не будут функционировать. В таком случае в клетках кишечного эпителия гены гемоглобина не работают не потому, что они там отсутствуют, а вследствие повреждения их структуры или выпадения незначительных по размерам последовательностей (типа ТАТА-бокса), регулирующих транскрипцию.
Экспериментальные данные, опровергнувшие и эту точку зрения на механизм дифференциации, были получены английским ученым Дж. Гердоном в начале 60-х годов (рисунок 1) на Xenopus laevis. Неоплодотворенные яйцеклетки облучали большой дозой ультрафиолета, которая инактивировала ядра, но практически не повреждала цитоплазму. С помощью микрохирургической техники в такие энуклеированные яйцеклетки пересаживались ядра из дифференцированных клеток — эпителия кишечника головастика. В некоторых случаях удалось получить полностью нормальных плодовитых взрослых особей.
Если для опыта брали ядра одной особи, то все развившиеся животные представляли собой клон, т. е. были сходны между собой так же, как однояйцевые близнецы человека. Из опытов Гердона можно сделать два вывода: во-первых, в процессе детерминации и дифференцировки не происходит необратимых повреждений генома; во-вторых, перенесение ядра тканевой клетки в неоплодотворенное яйцо по крайней мере в некоторых случаях приводит к полному возврату дифференцированного состояния и детерминации.
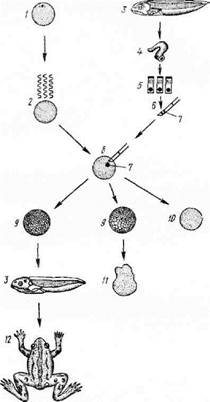
Рисунок 1. Клонирование Xenopus laevis - развитие взрослой особи из яйцеклетки, ядро которой заменено ядром из соматической клетки кишечного эпителия головастика:
1 — неоплодотворенное яйцо, 2 — УФ-облучение, 3 — головастик, 4 — кишечник головастика, 5— клетки кишечного эпителия, 6 — микропипетка, 7 — ядро эпителиальной клетки, 8 — яйцо-реципиент, 9 — бластула, 10 — неделяшаяся клетка, 11— ненормальный эмбрион, 12— взрослая лягушка
Другие примеры частичной обратимости дифференцированного состояния могут быть показаны с помощью гибридизации соматических клеток in vitro. В настоящее время техника подобных экспериментов достаточно высока, поэтому довольно легко можно получить гибриды между клетками даже далеких видов с разными типами молекулярной организации генома, например, между клетками птиц и млекопитающих. Если слить эритроциты птиц, ядра которых полностью потеряли генетическую активность, с клетками HeLa (человеческого происхождения), то в полученных гибридных клетках быстро активируются эритроцитарные ядра; в них синтезируются РНК, ДНК и белки, специфичные для данного вида птиц. Однако в природе едва ли существуют условия, при которых резко нарушается стабильность дифференцированного состояния, а тем более происходит передетерминация. Таким образом, поскольку детерминация и дифференцировка не связаны с количественными или качественными изменениями генома (в абсолютном большинстве случаев), то принято считать, что эти процессы основаны на эпигеномной наследственности. Сущность ее состоит в постоянном воспроизведении в ряду поколений соматических клеток такой надмолекулярной организации хромосом, которая позволяет функционировать в каждом типе клеток строго определенным наборам генов.
У высших растений геном соматических клеток также в основном репрессирован, и эта репрессия поддерживается эпигеномной наследственностью. Однако в этом случае полная дерепрессия генома в культуре растительной ткани наблюдается чаще. Например, из соматических клеток моркови и табака можно получить полноценные фертильные растения.
Механизм становления детерминации пока неизвестен, однако ясно, что у многих животных, например у амфибий, первичная детерминация связана с химической неоднородностью различных участков яйцеклетки. Поэтому и детерминация ядер, оказавшихся в ходе дробления в районе вегетативного полюса, будет иной, чем тех, которые попадут в цитоплазму анимального полюса.
Дата добавления: 2015-12-01; просмотров: 1596;