Рецессивный эпистаз у мышей
Ген А определяет синтез черного пигмента, ген В способствует распределению пигмента по длине волоса, рецессивный ген b не влияет на окраску шерсти. Рецессивный ген а нарушает синтез пигмента и в гомозиготном состоянии подавляет действие гена В (ааВ– альбиносы).
При скрещивании черных и белых мышей в F1 получаются лишь мыши типа агути (АаВв). В F2 9/16 мышей имеют окраску агути, 3/16 – черную и 4/16 – белую. Такое же расщепление характерно и для комплементарного взаимодействия генов.
Характерные признаки эпистатического взаимодействия генов:
1. действие двух пар генов на один признак;
2. подавление проявления гипостатичного гена в F1;
3. изменение формулы дигибридного расщепления в F2 за счет расширения доли особей с фенотипом гена супрессора, при этом характерные формулы расщепления для доминантного эпистаза 13:3, и 12:3:1, для рецессивного эпистаза – 9:3:4.
3. Полимерия – это тип неаллельного взаимодействия генов, при котором несколько пар неаллельных генов влияют на формирование одного признака, вызывая сходные изменения.
Явление полимерии было открыто в 1909 г. шведским генетиком Нильсоном-Эле, который описал серию однозначно действующих генов, определяющих окраску эндосперма зерна пшеницы.
Это случай так называемой кумулятивной полимерии (сложной) когда степень проявления признака зависит от числа доминантных аллелей в генотипе. Так наследуется, например, длина початка у кукурузы.
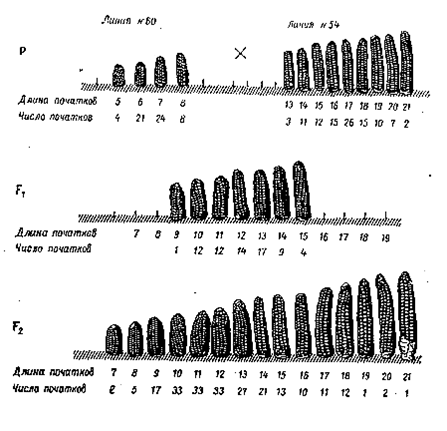
Наследование и изменчивость длины початков (в сантиметрах) у Zea mays в F1и F2
Одна из исходных линий (№ 60) имеет длину початков в пределах от 5 до 8 см, линия № 54 – от 13 до 21 см. Гибриды F1 имеют средние значения длины початков. Растения F2 фенотипически неоднородны, длина початков варьирует от 7 до 21 см. При этом длина початка пропорциональна числу (дозе) доминантных генов в генотипе.
По типу кумулятивной полимерии наследуется пигментация кожи у человека. Например, в потомстве у чернокожего мужчины и белой женщины (или наоборот) рождаются дети с промежуточным цветом кожи – мулаты. У супружеской пары мулатов рождаются дети с цветом кожи от черного до белого, что определяется числом доминантных аллелей в генотипе:
А1А1А2А2 × а1а1а2а2
 чернокожий белый
чернокожий белый
А1а1А2а2 × А1а1А2а2
мулаты
А1А1А2А2 А1А1А2а2 А1а1А2А2 А1а1А2а2 А1А1а2а2 а1а1А2А2 А1а1а2а2 а1а1А2а2 а1а1а2а2


 1/16 2/16 2/16 4/16 1/16 1/16 2/16 2/16 1/16
1/16 2/16 2/16 4/16 1/16 1/16 2/16 2/16 1/16
чернокожие «темные» мулаты «светлые» белые
При некумулятивной полимерии(простой), наличие в генотипе хотя бы одного доминантного аллеля полимерных генов определяет треугольную форму плодов. Например, при скрещивании растений пастушьей сумки с треугольными плодами (стручками) с растением с овальными плодами в F1 образуются растения с плодами треугольной формы.
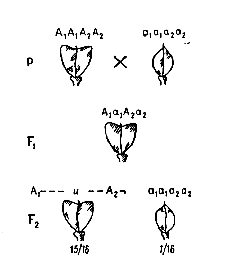
Наследование формы стручка у Capsella bursa pastoris при взаимодействии двух пар генов
При их самоопылении в F2 наблюдается расщепление на растения с треугольными и овальными плодами в соотношении 15:1. Если расщепление в F2 составляет 63:1, то в формировании признака участвуют 3 пары однозначных генов.
При полимерном типе наследования возможно проявление трансгрессий. Трансгрессия – форма, у которой степень проявления признака больше, чем у родительских форм.
Трансгрессии могут быть положительными и отрицательными:
P: A1A1a2a2A3A3a4a4×a1a1A2A2a3a3A4A4
F1: A1aA2a2A3a3A4a4
F2: A1A1A2A2A3A3A4A4 aa1a2a2a3a3a4a4
положительная отрицательная трансгрессии
Таким образом, трансгрессии проявляются в F2, когда родительские формы не обладают крайним проявлением признаков и не несут всех доминантных (при положительной трансгрессии) или всех рецессивных (при отрицательной трансгрессии) аллелей.
Гены-модификаторы – гены, усиливающие или ослабляющие действие основного гена.
Изучение окраски млекопитающих показало, что наряду с крайними формами, обладающими полным развитием пигмента (черная окраска) или его отсутствием (альбиносы), имеется целый ряд промежуточных форм – сероватых, бурых, желтых. Окраска шерсти зависит от наличия генов-модификаторов, не имеющих собственное проявление, но изменяющих действие основного гена.
Гены-модификаторы контролируют вкус, цвет и аромат плодов, поэтому их рекомендуется накапливать для улучшения признаков сортов плодовых культур.
2. Пенетрантность и экрессивность. Норма реакции. Плейотропный эффект гена.
Пенетрантность гена – это доля особей, у которых проявляется ожидаемый фенотип.
Экспрессивность – это степень выраженности фенотипа.
Многие гены имеют полную пенетрантность и экспрессивность. В опытах Менделя все горошины, несущие доминантный аллель определяющий желтую окраску были желтыми ( как в гомозиготном так и в гетерозиготном состоянии), а все горошины гомозиготные по аллелю определяющего зеленую окраску- зелеными. Все люди генотипа IА IА или IАi имеют группу крови А, люди генотипа IВ IВ или IВi группу крови В, а генотип IА IВ определяет группу крови АВ.
Примером неполной пенетрантности и экспрессивности может служить проявление доминантного гена, вызывающего хорею Гентингтона у человека. Люди несущие этот доминантный ген, заболевают в различном возрасте, некоторые из них остаются здоровыми в течении почти всей жизни. Болезнь начинается с непроизвольных, подергиваний головы, конечностей и туловища и прогрессируя, приводит к дегенеративным изменениям нервной системы, потере физических и умственных сил и смерти. Возраст первого проявления хореи Гентингтона – от младенчества до старости. Данный ген имеет неполную пенетрантность поскольку, известно что, у некоторых носителей он так и не проявляется и не заболевают. Этот ген имеет варьирующую экспрессивность: его носители заболевают в различном возрасте, т.е. он оказывает на их жизнь различное влияние. Причины, которые влияют на проявление гена у одних особей и не проявляется у других, могут быть влияние внешней среды и генотипа. Роль среды в проявлении гена очевидна и в ауксотрофных мутациях и других условных леталей. К примеру дрозофилы несущие некоторые температурочувствительные летальные мутации, обладают нормальной жизнеспособностью при 20 С, но при 29 С они теряют подвижность или погибают внешняя среда также оказывает влияние на экспрессивность морфологических признаков. Например, у человека полидактилия проявляется в присутствии доминантного гена D, который контролирует число лучистых костей. Экспрессивность генотипа Dd варьирует у одного и того же человека, который может иметь на одной руке пять пальцев, а на другой шесть. В этом случае варьирующая экспрессивность определяется внутренней средой развивающегося организма. На пенетрантность и экспрессивность гена также могут оказывать влияние другие гены данной особи, что выражено в случае генов-модификаторов и эпистатических генов.
Плейотропия – явление, которое заключается в том, что один ген оказывает влияние на несколько признаков.
Например, влияние плейотропного гена окраски меха у лисиц на жизнеспособность потомства.
Ген платиновой окраски является доминантным по отношению к серебристо-черной. Однако в гомозиготном состоянии он приводит к гибели зародышей (АА) на ранних стадиях:
Аа × Аа
Платиновые Платиновые
2Аа : 1аа
Платиновые Серебристо-черные
Выживают только платиновые лисицы, гетерозиготные по этому гену. По этой же схеме наследуется наличие (аа) и отсутствие (Аа) чешуи у зеркального карпа, серая (Аа) и черная (аа) окраска шерсти каракулевых овец.
У человека известен доминантный ген, определяющий признак «паучьи пальцы» (синдром Марфана). Одновременно этот же ген определяет аномалию хрусталика глаза и порок сердца.
Литература
1. Айала, Ф. Современная генетика / Ф. Айала, Дж. Кайгер. – М.: Мир, 1987. – Т.1. – 295 с; Т.2. – 368 с; Т.3.
2. Алиханян, С. И. Общая генетика / С. И. Алиханян, А. П. Акифьев,
Л. С. Чернин. – М.: Высш. шк., 1985.
3. Бокуть, С. Б. Молекулярная биология: молекулярные механизмы хранениия, воспроизведения и реализации генетической информации / С. Б. Бокуть, Н. В. Герасимович, А. А. Милютин. – Мн.:Высш. шк., 2005.
4. Дубинин, Н. П. Общая генетика / Н. П. Дубинин. – М.: Наука, 1986.
5. Жимулев, И. Ф. Общая и молекулярная генетика / И. Ф. Жимулев. – Новосибирск: Изд-во Новосибирского ун-та, 2002.
6. Жученко, А. А. Генетика / А. А Жученко, Ю. Л. Гужов,
В. А. Пухальский. – М.: Колос, 2004.
Дата добавления: 2015-12-01; просмотров: 1366;