Глава 17. Пути метаболизма глюкозы
Глюкоза является основным метаболитом и транспортной формой углеводов в организме человека и животных. Источниками глюкозы являются углеводы пищи, гликоген тканей и процесс глюконеогенеза в печени и корковом веществе почек. Для включения глюкозы в метаболизм она должна фосфорилироваться с образованием глюкозо‑6‑фосфата (Г‑6‑Ф), который далее может превращаться по различным метаболическим путям. На Рис. 17.1. представлены основные пути метаболизма глюкозы.
Гликолиз
Гликолиз – главный путь катаболизма глюкозы путем последовательных ферментативных превращений до лактата (без потребления кислорода – анаэробный гликолиз) или через окислительное декарбоксилирование пирувата до СО2 и Н2О (в присутствии кислорода – аэробный гликолиз).
Процесс аэробного гликолиза включает несколько стадий:
1. Аэробный гликолиз – процесс окисления глюкозы с образованием двух молекул пирувата;
2. Общий путь катаболизма, включающий окислительное декарбоксилирование пирувата до ацетил КоА и его дальнейшее окисление в цикле трикарбоновых кислот;
3. Цепь тканевого дыхания, сопряженная с реакциями дегидрирования, происходящими в процессе распада глюкозы.
Суммарный выход АТФ при окислении 1 моль глюкозы до СО2 и Н2О составляет 38 моль.
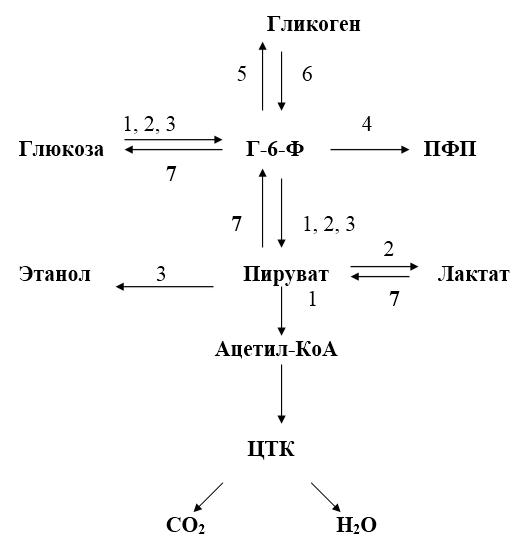
Рис. 17.‑1. Общая схема путей метаболизма глюкозы.
1 – аэробный гликолиз; 2 – анаэробный гликолиз; 3 – спиртовое брожение; 4 – пентозофосфатный путь; 5 – синтез гликогена; 6 – распад гликогена; 7 – глюконеогенез.
Анаэробный гликолиз – процесс расщепления глюкозы с образованием в качестве конечного продукта лактата. Этот процесс протекает без использования кислорода и поэтому не зависит от работы митохондриательной сети. АТФ здесь образуется за счет реакций субстратного фосфорилирования. Баланс АТФ при анаэробном гликолизе составляет 2 моль в расчете на 1 моль глюкозы.
Аэробный гликолиз происходит во многих органах и тканях и служит основным, хотя и не единственным, источником энергии для жизнедеятельности.
Кроме энергетической функции гликолиз может выполнять и анаболические функции. Метаболиты гликолиза используются для синтеза новых соединений. Так, фруктозо‑6‑фосфат и глицеральдегид‑3‑фосфат участвуют в образовании рибозо‑5‑фосфата – структурного компонента нуклеотидов. 3‑фосфоглицерат может включаться в синтез аминокислот, таких как серин, глицин, цистеин. В печени и жировой ткани ацетил‑КоА, образующийся из пирувата, используется как субстрат при биосинтезе жирных кислот, холестерина.
Анаэробный гликолиз активизируется в мышцах при интенсивной мышечной работе, происходит в эритроцитах (в них отсутствуют митохондрии), а также в разных условиях ограниченного снабжения их кислородом (спазм и тромбоз сосудов, формирование атеросклеротических бляшек).
Пентозофосфатный путь (ПФП)
ПФП, называемый также гексозомонофосфатным шунтом, служит альтернативным путем окисления глюкозо‑6‑фосфата. По ПФП в печени метаболизируется до 33 % всей глюкозы, в жировой ткани – до 20 %, в эритроцитах – до 10 %, в мышечной ткани – менее 1 %. Наиболее активно ПФП протекает в жировой ткани, печени, коре надпочечников, эритроцитах, молочной железе в период лактации, семенниках. ПФП состоит из 2 фаз (частей) – окислительной и неокислительной.
В окислительной фазе глюкозо‑6‑фосфат необратимо окисляется в пентозу – рибулозо‑5‑фосфат, и образуется восстановленный НАДФН2. В неокислительной фазе рибулозо‑5‑фосфат обратимо превращается в рибозо‑5‑фосфат, метаболиты гликолиза и другие фосфорилированные сахара.
Биологическая роль ПФП:
1. Наработка восстановленного НАДФН2 для восстановительных биосинтезов (жирных кислот, холестерина и т. д.).
2. Синтез пентозофосфатов для образования нуклеиновых кислот и некоторых коферментов.
3. Синтез моносахаридов с числом углеродных атомов от 3 до 8.
4. Обезвреживание ксенобиотиков – необходим НАДФН2.
5. В растениях – участие в темновой фазе фотосинтеза как акцептор СО2.
ПФП не приводит к синтезу АТФ, т. е. не выполняет энергетическую функцию.
Глюконеогенез (ГНГ)
Глюконеогенез – это синтез глюкозы из неуглеводных предшественников. Основной функцией ГНГ является поддержание уровня глюкозы в крови в период длительного голодания и интенсивных физических нагрузок. Процесс протекает в основном в печени и менее интенсивно в корковом веществе почек, а также в слизистой оболочке кишечника. Эти ткани могут обеспечивать синтез 80–100 г глюкозы в сутки.
Первичными субстратами (предшественниками) в ГНГ являются лактат, глицерол, большинство аминокислот. Включение этих субстратов в ГНГ зависит от физиологического состояния организма.
Лактат – продукт анаэробного гликолиза, образуется в работающих мышцах и, непрерывно в эритроцитах. Таким образом, лактат используется в ГНГ постоянно. Глицерол высвобождается при гидролизе жиров в жировой ткани в период голодания или при длительной физической нагрузке. Аминокислоты образуются в результате распада мышечных белков и выполняются в ГНГ при длительном голодании или продолжительной мышечной работе. Аминокислоты, которые при катаболизме превращается в пируват или метаболиты цикла трикарбоновых кислот, могут рассматриваться как потенциальные предшественники глюкозы и носят название гликогенных.
Из всех аминокислот, поступающих в печень, примерно 30 % приходится на долю аланина. Это объясняется тем, что при расщеплении мышечных белков образуются аминокислоты, многие из которых превращаются сразу в пируват или сначала в оксалоацетат, а затем в пируват. Последний превращается в аланин, приобретая аминогруппу от других аминокислот. Аланин из мышц переносится кровью в печень, где снова преобразуется в пируват, который частично окисляется и частично включается в ГНГ. Такая последовательность превращений приводит к формированию глюкозо‑аланинового цикла.
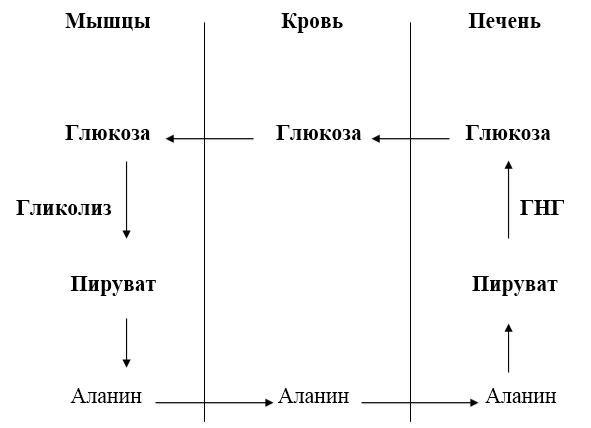
Рис. 17.2. Глюкозо‑аланиновый цикл.
Дата добавления: 2016-01-30; просмотров: 6034;