Мембранный транспорт
Транспорт веществ внутрь и наружу клетки, а также между цитоплазмой и различными субклеточными органеллами (митохондриями, ядром и т.д.) обеспечивается мембранами. Если бы мембраны были глухим барьером, то внутриклеточное пространство оказалось бы недоступным для питательных веществ, а продукты жизнедеятельности не могли бы быть удалены из клетки. В то же время при полной проницаемости было бы невозможно накопление определенных веществ в клетке. Транспортные свойства мембраны характеризуются полупроницаемостью: некоторые соединения могут проникать через нее, а другие - нет:
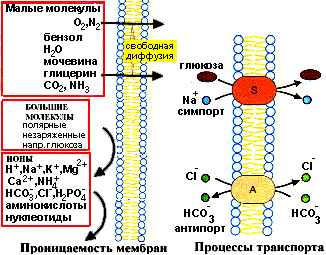
Проницаемость мембран для различных веществ
Одна из главных функций мембран - регуляция переноса веществ. Существуют два способа переноса веществ через мембрану: пассивный и активный транспорт:
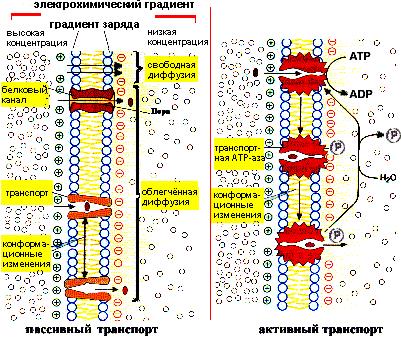
Транспорт веществ через мембраны
Пассивный транспорт. Если вещество движется через мембрану из области с высокой концентрацией в сторону низкой концентрации (т.е. по градиенту концентрации этого вещества) без затраты клеткой энергии, то такой транспорт называется пассивным, или диффузией. Различают два типа диффузии: простую и облегченную.
Простая диффузия характерна для небольших нейтральных молекул (H2O, CO2, O2), а также гидрофобных низкомолекулярных органических веществ. Эти молекулы могут проходить без какого-либо взаимодействия с мембранными белками через поры или каналы мембраны до тех пор, пока будет сохраняться градиент концентрации.
Облегченная диффузия. Характерна для гидрофильных молекул, которые переносятся через мембрану также по градиенту концентрации, но с помощью специальных мембранных белков - переносчиков. Для облегченной диффузии, в отличие от простой, характерна высокая избирательность, так как белок переносчик имеет центр связывания комплементарный транспортируемому веществу, и перенос сопровождается конформационными изменениями белка. Один из возможных механизмов облегченной диффузии может быть следующим: транспортный белок (транслоказа) связывает вещество, затем сближается с противоположной стороной мембраны, освобождает это вещество, принимает исходную конформацию и вновь готов выполнять транспортную функцию. Мало известно о том, как осуществляется передвижение самого белка. Другой возможный механизм переноса предполагает участие нескольких белков-переносчиков. В этом случае первоначально связанное соединение само переходит от одного белка к другому, последовательно связываясь то с одним, то с другим белком, пока не окажется на противоположной стороне мембраны.
Активный транспорт имеет место в том случае, когда перенос осуществляется против градиента концентрации. Такой перенос требует затраты энергии клеткой. Активный транспорт служит для накопления веществ внутри клетки. Источником энергии часто является АТР. Для активного транспорта кроме источника энергии необходимо участие мембранных белков. Одна из активных транспортных систем в клетке животных отвечает за перенос ионов Na+ и K+ через клеточную мембрану. Эта система называется Na+ - K+ - насос. Она отвечает за поддержание состава внутриклеточной среды, в которой концентрация К+ выше, чем Na+ :
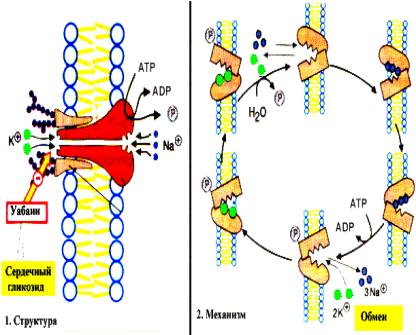
Механизм действия Na+, K+-АТР-азы
Градиент концентрации калия и натрия поддерживается путем переноса К+ внутрь клетки, а Na+ наружу. Оба транспорта происходят против градиента концентрации. Такое распределение ионов определяет содержание воды в клетках, возбудимость нервных клеток и клеток мышц и другие свойства нормальных клеток. Na+ ,K+ -насос представляет собой белок - транспортную АТР-азу. Молекула этого фермента является олигомером и пронизывает мембрану. За полный цикл работы насоса из клетки в межклеточное вещество переносится три иона Na+, а в обратном направлении - два иона К+. При этом используется энергия молекулы АТР. Существуют транспортные системы для переноса ионов кальция (Са2+ - АТР-азы), протонные насосы (Н+ - АТР-азы) и др. Симпорт это активный перенос вещества через мембрану, осуществляемый за счет энергии градиента концентрации другого вещества. Транспортная АТР-аза в этом случае имеет центры связывания для обоих веществ. Антипорт - это перемещение вещества против градиента своей концентрации. При этом другое вещество движется в противоположном направлении по градиенту своей концентрации. Симпорт и антипорт могут происходить при всасывании аминокислот из кишечника и реабсорбции глюкозы из первичной мочи. При этом используется энергия градиента концентрации ионов Na+, создаваемого Na+, K+-АТР-азой.
6)
К мембранным белкам относятся белки, которые встроены в клеточную мембрану или мембрану клеточной органеллы или ассоциированы с таковой. Около 25 % всех белков являются мембранными.[1]
Содержание
[показать]
Классификация[править | править вики-текст]
Мембранные белки могут быть классифицированы по топологическому или биохимическому принципу. Топологическая классификация основана на том, сколько раз белок пересекает липидный бислой. В соответсвии с этим критерием белки подразделяются намонотопические, битопические и политопические:
·монотопические белки взаимодействуют с одной поверхностью мембраны и не пересекаю её;
·битопические пронизывают мембрану насквозь и взаимодействуют с обеими её поверхностями;
·политопические пронизывают мембрану несколько раз (многократное взаимодействие с липидами).
Понятно, что первые относятся к периферическим белкам, а вторые и третьи к интегральным.
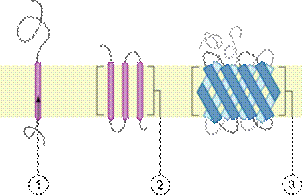
Различные категории политопических белков. Связывание с мембраной за счёт (1) единичной трансмембранной альфа-спирали, (2) множественных трансмембранных альфа-спиралей, (3) бета-складчатой структуры.
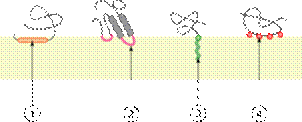
Различные категории интегральных монотопических белков. Связывание с мембраной за счёт (1) амфипатической альфа-спирали, параллельной плоскости мембраны, (2) гидрофобной петли, (3) ковалентно соединённогожирнокислотного остатка, (4) электростатического взаимодействия (прямого или кальций-опосредованного).
Топологическая классификация[править | править вики-текст]
По отношению к мембране мембранные белки делятся на поли- и монотопические.
·Политопические, или трансмембранные, белки полностью пронизывают мембрану и, таким образом, взаимодействуют с обеими сторонами липидного бислоя. Как правило, трансмембранный фрагмент белка является альфа-спиралью, состоящей из гидрофобных аминокислот (возможно от 1 до 20 таких фрагментов). Только у бактерий, а также вмитохондриях и хлоропластах трансмембранные фрагменты могут быть организованы как бета-складчатая структура(от 8 до 22 поворотов полипептидной цепи).
·Интегральные монотопические белки постоянно встроены в липидный бислой, но соединены с мембраной только на одной стороне, не проникая на противоположную сторону.
Биохимическая классификация[править | править вики-текст]
По биохимической классификации мембранные белки делятся на интегральные и периферические.
·Интегральные мембранные белки прочно встроены в мембрану и могут быть извлечены из липидного окружения только с помощью детергентов или неполярных растворителей. По отношению к липидному бислою интегральные белки могут быть трансмембранными политопическими или интегральными монотопическими.
·Периферические мембранные белки являются монотопическими белками. Они либо связаны слабыми связями с липидной мембраной, либо ассоциируют с интегральными белками за счёт гидрофобных, электростатических или других нековалентных сил. Таким образом, в отличие от интегральных белков они диссоциируют от мембраны при обработке соответствующим водным раствором (например, с низким или высоким pH, с высокой концентрацией соли или под действием хаотропного агента). Эта диссоциация не требует разрушения мембраны.
Мембранные белки могут быть встроены в мембрану за счёт жирнокислотных или пренильных остатков либогликозилфосфатидилинозитола, присоединённых к белку в процессе их посттрансляционной модификации.
7)Углеводная часть гликолипидов и гликопротеинов плазматической мембраны всегда находится на наружной поверхности мембраны, контактируя с межклеточным веществом. Углеводы плазматической мембраны выполняют роль специфических лигандов для белков. Они образуют участки узнавания, к которым присоединяются определенные белки; присоединившийся белок может изменить функциональное состояние клетки.
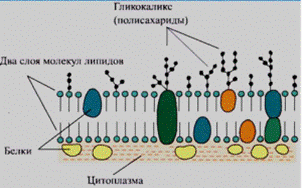
Функции углеводов.
В наружной мембране эритроцитов некоторые полисахариды содержат N-аце-тилнейраминовую кислоту на концах цепей. Если эритроциты выделить из крови, обработать in vitro нейраминидазой, отщепляющей N-ацетилнейраминовую кислоту от мембранных углеводов, и вновь ввести в кровь тому же животному, то обнаруживается, что время полужизни таких эритроцитов в крови уменьшается в несколько раз: они задерживаются в селезенке и разрушаются. Как выяснилось, в клетках селезенки есть рецептор, узнающий углевод, который утратил концевые остатки нейраминовой кислоты. Возможно, что такой механизм обеспечивает отбор селезенкой «состарившихся» эритроцитов и их разрушение.
Известно, что в суспензии клеток, выделенных из какой-либо ткани, через некоторое время образуются агрегаты клеток, причем в каждом агрегате, как правило, оказываются клетки одного типа. Например, в суспензии клеток, полученных из гаструлы, образуется три вида агрегатов: каждый из них содержит клетки, принадлежащие одному и тому же зародышевому листку — эктодерме, мезодерме или эндодерме. Узнавание между клетками обеспечивается, в частности, взаимодействием мембранных углеводов одной клетки с белками-рецепторами другой клетки (рис. 9.39). Эти механизмы узнавания могут участвовать в таких процессах, как гистогенез и морфогенез. Однако есть и другие механизмы, обеспечивающие межклеточные контакты.
Полисахариды клеточной мембраны наряду с белками выполняют роль антигенов при развитии клеточного иммунитета, в том числе при реакции отторжения трансплантата. Они также служат местами узнавания при заражении патогенными вирусами и микроорганизмами. Например, вирус гриппа при проникновении в клетку сначала присоединяется к ее мембране, взаимодействуя с полисахаридом определенной структуры.
8)леточные мембраны обладают избирательной проницаемостью: через них медленно диффундируют глюкоза, аминокислоты, жирные кислоты, глицерол и ионы, причем сами мембраны в известной мере активно регулируют этот процесс — одни вещества пропускают, а другие нет. Существует четыре основных механизма для поступления веществ в клетку или вывода их из клетки наружу: диффузия, осмос, активный транспорт и экзо- или эндоцитоз. Два первых процесса носят пассивный характер, то есть не требуют затрат энергии; два последних — активные процессы, связанные с потреблением энергии.
Избирательная проницаемость мембраны при пассивном транспорте обусловлена специальными каналами — интегральными белками. Они пронизывают мембрану насквозь, образовывая своего рода проход. Для элементов K, Na и Cl есть свои каналы. Относительно градиента концентрации молекулы этих элементов движутся в клетку и из неё. При раздражении каналы натриевых ионов раскрываются, и происходит резкое поступление в клетку ионов натрия. При этом происходит дисбаланс мембранного потенциала. После чего мембранный потенциал восстанавливается. Каналы калия всегда открыты, через них в клетку медленно попадают ионы калия
8)

Дата добавления: 2016-01-30; просмотров: 5533;