Почему жизнь дискретна
О дискретности жизни и смене поколений образно сказал А. С. Пушкин в «Евгении Онегине»:
«Увы! на жизненных браздах
Мгновенной жатвой поколенья,
По тайной воле провиденья,
Восходят, зреют и падут;
Другие им вослед идут...»
Можно сказать, что естествознание нашего века началось с осознания дискретности мира. В физике «сплошная, гладкая» материя – континуум, с непрерывными превращениями, сменилась на нечто дискретное в пространстве и времени, с квантованными превращениями. В биологии представления о слитой наследственности, многим казавшиеся самыми естественными, с 1900 г., после «переоткрытия» законов Менделя, стали интенсивно вытесняться концепцией дискретного наследования.
Дискретность организменного уровня не требовала доказательств: настолько она очевидна, достаточно взглянуть на самих себя. Она и легла в основу эволюционных представлений об изменении во времени, эволюции форм. Дискретность на клеточном уровне была доказана экспериментально с применением микроскопов; дискретность видов лежала в основе дарвинизма; пространственная отграниченность и дискретность локальных экосистем активно изучаются в настоящее время. Итак, всю биологию пронизывает идея дискретности. И все же... И все же почему не может существовать непрерывная живая плазма, вечная во времени (типа живого мыслящего океана С. Лема)? Почему мы должны рождаться и умирать, сменяя друг друга в поколениях, а наши внуки должны нас «вытеснять из мира»?
Этот вопрос «почему» для традиционной биологии непрост, а для субстратного подхода, опирающегося на неопределенную и рыхлую идею «саморазвития», он во многих случаях просто убийственный. Действительно, зачем «саморазвивающейся» системе рвать себя на куски, отмирать, теряя при этом почти все? Разве не хватило бы простой «косметики», сохраняющей непрерывность структур? Посмотрим, как можно объяснить дискретность с позиций С + Э подхода, рассмотрев ее на разных уровнях биологической организации.
Обратимся к клеточному уровню. Для этого вернемся к самому началу, к происхождению жизни и появлению фазово‑обособленных частиц в абиогенном органическом бульоне, образовавшемся под влиянием накачки энергией. Отметим: в фазово‑обособленных, т. е. дискретных , единицах резко ускоряются процессы переноса энергии (электронов) и под их действием начинают работать первые циклы вещества. Таким образом, дискретность усиливает взаимодействие: энергия – вещество, ускоряя его при многократном использовании вещества, которое без циклов быстро израсходовалось бы, в то время как накачка энергией со стороны может идти вечно. Законы здесь только физико‑химические, ни о каком «саморазвитии» нет смысла говорить (эволюцию клеток мы обсуждали подробно в гл. 7 и 8).
Существует ли оптимальный размер клеток? Или: почему клетке выгодно быть определенной величины? Нижний предел ясен – он определяется минимальными размерами, при которых возможно самостоятельное существование, т. е. метаболизм и воспроизводство. А каковы верхние границы? Для определения верхних размеров издавна использовался энергетический подход. С прошлого века известны правила соотношения поверхности и объема клеток, получившие название законов Рубнера. Суть рассуждений сводится к тому, что с увеличением размеров клетки энергетическая эффективность ее функционирования падает. Это очевидно из самых простых соображений: расход энергии на рост (метаболизм) клетки пропорционален массе клетки или ее объему, а приток энергии пропорционален ее поверхности, так как питание она получает через поверхность. Если представить клетку в виде шарика, то отношение поверхности к объему шарика падает с ростом радиуса по обратному закону. Следовательно, масса (m ) клетки изменяется во времени таким образом:
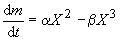
где α и β – константы питания и метаболизма. Остановке роста соответствует 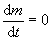
. Отсюда максимальный X (радиус клетки‑шарика) определяется в виде 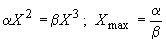
т. е. верхний размер определяется соотношением констант питания и метаболизма. Константа питания зависит как от внутренних характеристик, так и от внешнего потока энергии.
В самом общем виде с увеличением потока энергии константа питания возрастает вначале линейно, затем по гиперболической кривой с насыщением, а дальнейшее увеличение потока энергии может оказаться вредным для клетки. Эти качественные рассуждения хорошо демонстрируют идею дискретности на клеточном, а вообще говоря, и на организменном уровнях. Они применимы для определения максимальных размеров представителей двух основных типов живых организмов: растений и животных.
Известно, что растения, независимо от природы, растут сначала быстро, затем рост постепенно замедляется и, наконец, прекращается совсем. Интуитивно ясно, что с увеличением размеров растения увеличивается приток энергии благодаря фотосинтезу, но зато увеличиваются трудности, связанные с переносом питательных веществ, особенно с подъемом неорганических солей от корней к листьям. В конечном счете притока энергии перестает хватать для покрытия расходов, и дерево останавливается в росте. Несложные расчеты показывают, что предельное значение высоты дерева определяется энергетическими константами фотосинтеза, фотодыхания и транспорта.
Аналогичны рассуждения о предельных значениях размеров животных. Расходы энергии у животного связаны с основным обменом (аналог фотодыхания), с перемещением тела (аналог переноса вещества в растении) и с ростом. Форма уравнения для роста животного соответствует таковой для растения или отдельной клетки. Могут получаться более сложные зависимости максимального размера от концентрации корма или интенсивности энергетического потока, но дискретность везде имеет место.
Для оценки размеров водных организмов особый интерес представляет сопоставление размеров пищи и типа питания (фильтрация или активное хватание) с учетом энергетики. Из самых общих соображений понятно, что должна существовать размерная граница между пассивной фильтрацией и активным поиском и захватом пищи; очень мелкие частицы энергетически невыгодно разыскивать и хватать, так как полученная энергия не компенсирует ее расхода. Действительно, по различным оценкам, самые мелкие частицы (до 0,1–1 мкм) отфильтровываются животными, образующими слизистую сеть, через которую движениями ресничек прогоняется вода; частицы более 50 мкм преимущественно потребляются животными‑хватателями, а промежуточные размеры соответствуют обоим типам питания. При этом соблюдается соответствие размеров животного‑хищника и его жертвы. Пример с фильтрующими китообразными, по форме не укладывающийся в изложенную схему, хорошо подтверждает неуниверсальность С + Э подхода без учета информационных аспектов. Хорошо известна большая роль информации у этих высокоорганизованных высших животных (млекопитающих). Они буквально «пасут» стаи криля – их основной пищи, мигрируя за ними, имея громадный выигрыш прежде всего по энергетике, связанной с поиском и перемещением. Само появление хищничества оказалось возможным, начиная лишь с определенного нижнего размера тела животных, так как оно требует избыточного расхода энергии на поиск, преследование и захват жертвы. С энергетикой связано и соотношение размеров прокариотных и позднее появившихся, в сотни раз более крупных, эукариотных клеток.
Дискретность более высоких уровней тоже определяется функциональными отношениями в системе: «живое – неживое», среди которых энергетические потоки играют существенную роль. Для популяционного уровня, например, широко распространены представления об оптимальном размере популяции. Причем ограничения на численность популяции сверху непосредственно связываются с ограничением по ресурсам: на этом основано одно из наиболее известных уравнений математической экологии – логистическое. Нижний критический уровень для популяции (если он перейден сверху вниз, то популяция вымирает) определяется разнообразными причинами, но он существует и для микробных популяций, и для популяций высших организмов. В литературе представление об оптимальных размерах популяций связывается с «принципом Олли».
Для чего нужна дискретность на следующем видовом уровне? Почему она отобралась в эволюции, так как нескрещиваемость видов приводит к вымиранию целых таксонов, среди которых было немало находок, безвозвратно потерянных живой природой? И здесь можно оценить энергетическую невыгодность полного скрещивания на генном уровне из‑за усложнения структуры генома и энергетической дороговизны ее содержания (см. гл. 6).
Вопрос о дискретности экосистем (биогеоценозов, биоакваценозов) хотя и дискуссионен, но, по‑видимому, тоже очевиден. Дискретность этого уровня наглядна с точки зрения структуры: мы видим хорошо выраженные типы растительности, животного мира, относящиеся к определенным регионам. Границы определяются географическими особенностями, пространственным положением, а также зависят от истории развития биоты в данном месте. Нас более интересует функциональный аспект, тесно связанный с энергетикой. По функции соседние экосистемы именно тем и отличаются, что потоки энергии и круговороты вещества в них гораздо интенсивнее, чем обмены между ними. Казалось бы, дискретность экосистемам ни к чему, по крайней мере с точки зрения структурного содержания.
Однако дискретность экосистем, а с ними и биотического круговорота ярко выражена не столько в экологическом, сколько в эволюционном плане. При длительном функционировании круговорота в каком‑либо месте, как мы знаем с позиций энергетики, должна происходить подгонка составных элементов круговорота в двух основных направлениях: в количественном, т. е. в увеличении потока захваченной энергии, и в качественном, т. е. в ускорении циклов лимитирующего вещества, в том числе и путем упрощения структур. И здесь мы можем видеть удивительные примеры согласованной эволюции «соучастников». Просто перечислим некоторые примеры, характеризующие разные трофические уровни: растения и их ризосфера, специфическая в каждом случае, особо интересен здесь симбиоз азотфиксирующих бактерий и растений типа бобовых; цветковые растения и насекомые‑опылители; многоярусные леса; птицы, обитающие в каждом ярусе таких лесов; и т. д. Возможно, одним из самых забавных вариантов такой подгонки может служить «запрещенный» по прямой энергетике случай животного с зеленой, фотосинтезирующей поверхностью. Это ленивец, в шерсти которого во влажном тропическом лесу размножаются водоросли. Они не дают ему питания (и здесь все сходится по энергетике с расчетами), но зато маскируют его: зеленоватый оттенок спасает малоподвижного животного от крупных хищных птиц.
Итак, дискретность всех уровней биологической организации выгодна прежде всего с энергетических позиций. Поэтому для понимания ее необходимости в развитии и эволюции живого достаточно привлечь С + Э подход. Для строгих количественных оценок дискретности структур (их старения и замен) нужно привлекать и информационные аспекты, т. е. использовать в комплексе С + Э + И подход.
Дата добавления: 2016-01-26; просмотров: 1309;