Динамические процессы в экосистеме
В экосистеме постоянно осуществляются процессы перехода веществ и энергии с одних уровней трофической цепи на другие, поэтому экосистема не является абсолютно стабильным неизменным образованием. В любой экосистеме постоянно происходит изменение численности и плотности популяций в результате взаимодействия хищников с жертвами, а жертв с источниками их питания. Но одновременно с этим подвижность экосистемы является относительной, например экосистемы таежных лесов или целинных степей существуют длительное время (сотни лет) и, на первый взгляд, стабильны, устойчивы, неизменны. За короткий отрезок времени в них трудно обнаружить значительные изменения в составе популяций или режимах факторов неживой природы, хотя в отдельных случаях массовые размножения некоторых видов животных (например, лесных насекомых) существенно изменяют экосистему на тот или иной отрезок времени, а иногда способствуют ее замене на другую.
Следовательно, экосистемы с одной стороны, действительно являются стабильными образованиями, а с другой стороны динамичны и подвижны в пространстве и времени.
В том случае, если бы экосистемы существовали короткий отрезок времени, и происходило бы быстрое замещение одной экосистемы на другую, то накопление питательных веществ и стабильное развитие и размножение живых организмов было бы невозможно. Подвижно-стабильное состояние биогеоценозов (экосистем) во времени и пространстве представляет собой результат двух процессов: гомеостаза и сукцессии.
Между всеми компонентами экосистемы (биогеоценоза) происходят процессы обмена веществом и энергией, эти процессы сбалансированы между собой, а сам биогеоценоз характеризуется устойчивостью. Таким образом для биогеоценоза свойственно состояние подвижного равновесия, или гомеостаза (от греч. homоs - тот же, подобный, stasis - состояние).
Гомеостаз с точки зрения кибернетики обеспечивается механизмами обратной связи. Принцип обратной связи состоит в том, что некоторый управляющий компонент той или иной системы получат данные (информацию) от управляемых компонентов и использует эту информацию для внесения коррективов в дальнейший процесс управления.
Действия механизма обратной связи можно рассмотреть на примере популяций двух видов: оленя и волка (рис.3.8). В данном случае волк является хищником и поедает оленей, а олень является жертвой. В случае, если численность популяции оленей начнет возрастать, хищник, питающийся только этой жертвой, имеет возможность увеличить свою численность, и соответственно качественно и количественно улучшить свою популяцию. В данном случае проявляется положительная обратная связь (рис. 3.8.).
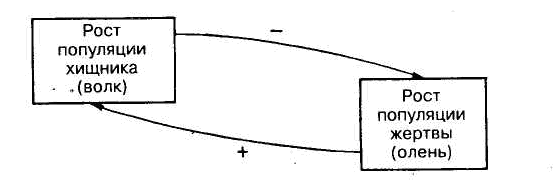
Рис.3.8 Пример действия механизма обратной связи
Но, питаясь оленями, в свою очередь волк (хищник) снижает численность популяции оленя. В данном случае проявляется отрицательная обратная связь.
Если численность волка превысит некоторый предел, то он соответственно снизит численность оленя и в итоге окажется перед необходимостью ограничения собственной численности из-за нехватки пищи, связанной с затрудненностью ее добычи. Аналогичная ситуация складывается с насекомыми, питающимися листьями деревьев. Пока имеется избыточная биомасса листьев, насекомые могут размножаться (положительная связь). Но резкое возрастание численности насекомых приведет к уменьшению биомассы листьев (отрицательная обратная связь) и вызовет гибель самих насекомых из-за недостатка пищи.
Таким образом, в биогеоценозе все время сохраняется равновесие, предотвращающее необратимое исчезновение тех или иных звеньев в пищевых цепях. Вследствие длительного эволюционного процесса названного Дарвином естественным отбором, численность оленя и волка всегда будет поддерживаться на определенно уровне. Любая экосистема всегда сбалансирована, устойчива (гомеостатична).
Пример, представлены на рис. 3.8 можно дополнить, введя в него абиотические факторы окружающей среды, которые с одной стороны независимо влияют на все звенья трофической цепи. В основерассмотренных процессов лежит принцип (закон) Ле Шателье (1884). Изменение внешних условий (температуры, давления) физико-химической равновесной системы вызывает в ней реакции, противодействующие производимому изменению [1,5].
Гомеостаз биогеоценоза не является застывшим состоянием. Биогеоценоз в сущности находится в стабильно-подвижном состоянии, поскольку в любой экосистеме идут процессы, меняющие ее во времени и пространстве. При этом происходят изменения в составе популяций, структуре экоистеме и ее продуктивности.
Рассмотренные выше процессы представляют собой обмен веществом и энергией между отдельными компонентами экосистемы, т.е. обмен информацией, передача которой производится по каналам обратной связи.
В ряде случаев передача информации может быть нарушена в связи с некоторыми обстоятельствами. Например, на оленей может начать охотиться еще один хищник, что может помешать волку, или внутри популяции волка может начаться инфекционная болезнь. В результате нарушается сбалансированность системы, этот процесс может быть обратимым или необратимым. С точки зрения кибернетики , в каналах обратной связи в данном случае, между популяциями хищника и жертвы возникли «помехи» или «шумы».
В качестве помех могут выступать и факторы неживой природы, например климатичесие условия. В результате засухи может сократиться биомасса зеленых растений, а следовательно уменьшится кормовая база для популяции оленей, что одновременно скажется и на популяции хищника (волка).
Влияние помех на популяцию избирательно и случайно, в результате их воздействия более стойкие особи выживают и передают наследственную информацию своим потомкам, а более слабые, для которых помехи оказались непреодолимыми погибают или не дают потомства. Таким образом, помехи выступают в качестве полезного фактора эволюции, поскольку под их воздействием осуществляется естественный отбор.
В процессе хозяйственной деятельности общество постоянно вмешивается в процессы, происходящие в экосистеме, за счет чего влияет как на отдельные ее компоненты, так и на экосистему в целом. Загрязнение объектов окружающей среды, изъятие природных ресурсов и прочие факторы не всегда приводят к исчезновению экосистемы или нарушению ее стабильности, но их воздействие не может быть безграничным. Экосистема может прекратить свое существование в случае, если механизм отрицательной обратной связи не сможет компенсировать отклонений, вызванных положительной обратной связью (появление другого хищника, резкое сокращение численности популяции в результате болезни).
Та область, в пределах которой механизмы отрицательной обратной связи способны, несмотря на стрессовые воздействия, сохранить устойчивость системы, хотя и в измененном виде, называют гомеостатическим плато (рис. 3.9).
Нарушение гомеостатического равновесия в результате резких антропогенных или естественных воздействий наиболее характерно для искусственных экосистем, например объектов сельского хозяйства. В ходе сельскохозяйственной деятельности регулирующую функцию выполняет человек. Например обеспечивает защиту возделываемых культур от насекомых-вредителей.
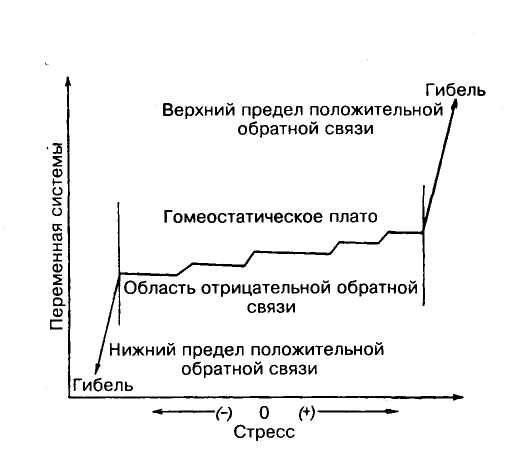
Рис.3.9. Схема формирования гомеостатического плато в экосистеме, в пределах которого посредством отрицательной обратной связи поддерживается относительная стабильность системы при воздействиях, вызывающих нарушение сбалансированности [1]
Последовательная смена биоценозов, преемственно возникающих на одной и той же территории в результате влияния природных факторов (в том числе внутренних противоречий развития самих биоценозов) или воздействия человека, называется сукцессией (от лат. succesio- последование, следую).
В данном случае действует принцип сукцессионного замещения. Природные биотические сообщества последовательно формируют закономерный ряд экосистем, ведущий к наиболее устойчивому в данных условиях состоянию климакса. Сукцессия - постепенный процесс изменения структуры и состава биоценоза.
В определенный отрезок времени, в который происходит экологическая сукцессия, изменяется абиотическая среда и видовая структура сообщества. Кульминацией развития сообщества является возникновение стабилизированной системы, называемой климаксом. В этом состояний система находится тогда, когда в ней на единицу энергии приходится максимальная биомасса и максимальное количество симбиотических связей между организмами (Ю. Одум, 1975) [2,6]. Однако к климаксу система приходит через ряд стадий развития, первые из которых часто называют стадией первых поселенцев. Поэтому, в более узком смысле, сукцессия - это последовательность сообществ, сменяющих друг друга в данном районе.
Сукцессия возникает на свободном пространстве и в зависимости от первоначального состояния субстрата выделяют первичную и вторичную сукцессию. Первичная сукцессия формируется на первоначально свободном субстрате, т.е. в месте абсолютно лишенном жизни (например, район отступления ледника). Вторичная сукцессияформируется на местах разрушенных сообществ, где сохраняется почва и часть организмов, т.е. на месте старого сообщества формируется новое, для которого данные абиотические условия являются более благоприятными. Например, после вырубки леса на опустевшей территории формируется экосистем луга, со временем на этой территории появляются кустарники, затем лиственные деревья, а после хвойные.
В ходе сукцессии увеличивается продуктивность биоценоза, т.е. синтез органического вещества на единицу площади, увеличивается видовое разнообразие.
В случае формирования первично сукцессии на свободном субстрате возникает растительность за счет переноса ветром спор и семян растений, либо за счет вегетативных органов оставшихся по соседству растений. Зарастание еловым лесом северных территорий России является примером первичной сукцессии.
В климатических условиях Севера ельник является последней стадией развития экосистемы, коренным биоценозом. Сперва же формируются березняки, ольховники, осинники, под пологом которых растут ели. Постепенно они перерастают березу и вытесняют ее, захватывая пространство (рис. 3.10).
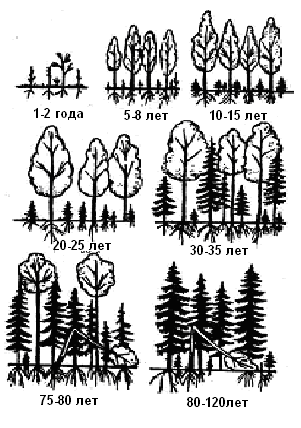
Рис. 3.10. Смена березняка ельником
Семена ели и березы легко переносятся ветром, но рост березы существенно превосходит скорость рота ели. К 6-10 годам ель едва достигает 50-60 см, а береза – 8-10 м. Однако сомкнувшиеся кроны берез обеспечивают возникновение особого микроклимата, за счет лиственного опада изменяется состав почвы, изменяется состав популяций живых организмов, формируется разнообразный травянистый покров, создаются консорции березы с окружающей средой. Таки образом, в благоприятном микроклимате рост елей продолжается и в конечном итоге береза не выдерживает конуренцию с ней за пространство и свет и вытесняется.
Очень часто вторичная сукцессия является результатом человеческой деятельности. Приведенный выше пример формирования елового леса в основном происходит в результате вторичной сукцессии, формирующейся на вырубках ранее существовавшего ельника.
Вторичная сукцессия заканчивается стабильной стадией сообщества через 150-250 лет, а первичная длится 1000 лет.
Примером антропогенной вторичной сукцессии может служить процесс эвтрофикации («цветения») водоемов. В результате антропогенной деятельности в водоемы, в частности искусственные водохранилища, поступают биогенные компоненты (чаще всего фосфор, реже азот, иногда углерод и кремний). Главная роль в этом процессе принадлежит фосфору, который является источником питательных веществ для водорослей, прежде всего сине-зеленых. Многие из них способны фиксировать молекулярный азот из атмосферы, тем самым снижая лимитирующее действие азота, а некоторые могут освобождать фосфор из продуктов метаболизма различных водорослей. Постепенно происходит зарастание водоема и массовая гибель рыб и других аэробных организмов из-за нехватки кислорода в воде, за счет чего биоценоз полностью изменяется.
Изучение специфики сукцессионных стадий позволяет оценить возможность изъятия и использования ресурсов природных систем, а следовательно дает возможность минимизировать негативное воздействие хозяйственной деятельности на окружающую среду.
Примеры экосистем
Все существующие экосистемы в первую очередь можно разделить на две большие группы: естественные (природные) и созданные человеком (антропогенные).
К антропогенным экосистемам можно отнести города, агроэкосисемы, космические корабли.
В составе природных экосистем можно выделить наземные, пресноводные и морские
Климатически обусловленные крупные совокупности наземных экосистем называются биомами.
Основные типы наземных биомов: пустынные, травянистые, лесные.
Пресноводные экосистемы классифицируются в зависимости от скорости циркуляции воды: лотические (лат. lotus - омывающий) - текучие воды (реки, ручьи) и лентические (лат. lente - медленно, спокойно) - стоячие воды (озера, пруды, лужи).
Классификация морских экосистем производится в засисимости от глубины водоема и вертикального размещения организмов: литоральная зона (лат. litoralis - прибрежный); батиальная зона (лат. bathis - глубокий); абиссиальная область (гр. abyssos - бездна).
По размерам экосистемы подразделяются на: микроэкосистемы (например, ствол упавшего дерева или поляна в лесу), мезоэкосистемы (лесной массив или степной колок), макроэкосистемы (тайга, море), экосистемы глобального уровня (планеты Земля).
Класифицировать экосистем также возможно по типу источника энергии
1. движимые Солнцем, малосубсидируемые
2. движимые Солнцем, субсидируемые другими естественными источниками
3. движимые Солнцем и субсидируемые человеком
4. движимые топливом.
В большинстве случаев могут использоваться и два источника энергии - Солнце и топливо.
К природным экосистемам, движимым Солнцем (малосубсидируемым) относятся открытые океаны и высокогорные леса. Источником энергии для данных экосистем практически является только Солнце, в связи с чем продуктивность их не велика. Обитатели таких экосистем адаптированы к небольшому количеству энергии и ресурсов, поэтому используют их эффективно. Малосубсидируемые экосистемы играют значительную роль в биосфере планеты, поскольку занимают огромные площади, например, океан
покрывает около 70 % поверхности земного шара. Таким образом, это основные системы жизнеобеспечения, механизмы, стабилизирующие и поддерживающие условия на планете. В процессе функционирования этих экосистем формируются климатические словия, очищаются существенные объемы атмосфеного воздуха, осуществляется круговорот воды, поддерживается температура и т.п., т.е. выполняются функции поддержания жизни на Земле. В независимости от человеческой деятельности здесь производится некоторое количество питательных веществ и других ресурсов. Кроме того, эти экосистемы обладают огромной эстетической ценностью.
Природные экосистемы, движимые Солнцем, субсидируемые другими естественными источниками. Этот тип экосистем обладает естественной плодородностью и производит излишки органического вещества, которые могут накапливаться. Кроме энергии солнечного излучения они получают энергию приливов, течений, прибоя, минеральные и органические вещества, поступающие с площади водосбора с дождем и ветром.
В качестве примеров таких экосистем можно привести дождевые леса, характеризующиеся избыточным плодородием, прибрежная часть эстуария типа Невской губы так же более плодородна, чем прилегающие участки суши, получающие то же количество солнечной энергии.
К экосистемам, движемым Солнцем и субсидируемым человеком отностся агроэкосистемы, как наземные, так и водные. Они получают одновременно энергию солнца и энергетические дотации от человека. В данном случае продуктиность в экосистемах поддерживается за счет человеческого труда (мышечной энергии) и энергии толива, которые используются на возделывание, орошение, удобрение, селекцию, переработку, транспортировку и т.п. Возделывание посевов кукурузы, картофеля, пшеницы и ржи не обходится без использования топлива. Самое продуктивное сельское хозяйство получает энергии примерно столько же, сколько самые продуктивные природные экосистемы второго типа.
Однако если в агроэкосистеме вся энергия направляется на производство продуктов питания определенного вида, то природа распределяет их между многими видами и накапливает на «черный день». Эта стратегия называется «стратегией повышения разнообразия в целях выживания».
В индустриально-городских экосистемах, движимых топливом энергия топлива полностью заменяет солнечную энергию. Продукт, произведенный экосистемами, движимыми Солнцем в городскую экосистему поставляют извне. Потребность плотно населенных городских районов в энергии на два-три порядка выше, чем в первых трех типах экосистем. Потребление энергии человеком в городе в среднем составляет более 80 млн ккал*год-1; для питания ему требуется всего около 1 млн ккал*год-1, следовательно, на все другие виды деятельности (домашнее хозяйство, транспорт, промышленность и т. д.) человек расходует в 80 раз больше энергии, чем требуется для физиологического функционирования организма. Разумеется, в развивающихся странах положение несколько иное.
Дата добавления: 2016-01-20; просмотров: 1333;