Межклеточные взаимодействия.
Нейроны, в соответствии со своими функциями, включены в систему связей, осуществляемых разными видами контактов.
Концевые отделы нервных волокон называются нервными окончаниями. Они служат для межклеточных взаимодействий и бывают рецепторными, межнейронными и нейроэффекторными.
Рецепторы – высокоспециализированные чувствительные образования, воспринимающие информацию из окружающей среды и преобразующие ее в нервный импульс. Различают экстерорецепторы – воспринимают информацию из внешней среды и интерорецепторы - воспринимают информацию из внутренней среды организма. По характеру воспринимаемых стимулов выделяют механо-, термо-, хемо- и фоторецепторы.
Рецепторные клетки нервного происхождения называют первичночувствующими рецепторами (термо-, хемо-, механо и фоторецепторы). Они имеют аксон и дендрит, преобразованный в воспринимающий элемент рецептора. Вторичночувствующие рецепторы – высокоспециализированные клетки ненервного происхождения (рецепторы вкуса, слуха и равновесия). Они воспринимают стимул и образуют особый контакт с дендритом чувствительного нейрона.
Межнейрональные контакты разделяют на неспециализированные и специализированные или синапсы. К неспециализированным контактам относят плотные или щелевые контакты и десмосомы. Плотные контакты представляют собой простое прилегание двух клеток с очень маленьким, порядка 3 – 5 мкм, расстоянием между их мембранами. Таких контактов много в коре больших полушарий головного мозга, сетчатке глаза, в стволе мозга. Плотные контакты обеспечивают химическую изоляцию, так как столь малое расстояние между нейронами препятствует диффузии высокомолекулярных веществ. Десмосомы – контакты механического типа. Между нейронами в ЦНС человека встречаются редко, более характерны для глиальных клеток.
Специализированные межнейронные контакты называются синапсами. Понятие ввел в 1906 году английский физиолог Шеррингтон.Синапс – (от греческого synapsis - соприкосновение, связь) - место контакта двух нейронов или нейрона и мышцы.
В зависимости от механизма передачи нервного импульса различают химические и электрические синапсы.
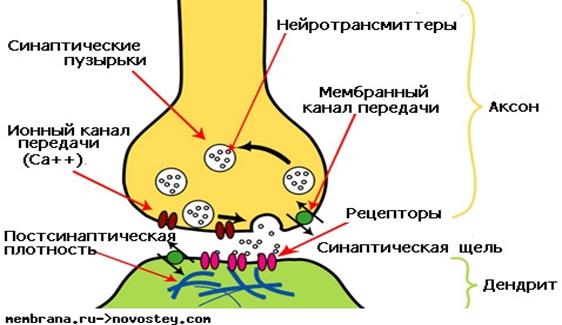
Рис. 1. Строение химического синапса
В составе любого синапса различают три главных компонента: пресинаптическую мембрану (пресинапс), синаптическую щель и постсинаптическую мембрану (постсинапс). Пресинапс, в большинстве случаев образован булавовидным расширением окончания аксона передающей клетки, постсинапс - представлен контактирующим участком мембраны воспринимающей клетки. Характерная особенность пресинаптической области химического синапса – наличие большого количества синаптических пузырьков. В пузырьках находится медиатор (нейротрансмиттер). При поступлении нервного импульса происходит высвобождение медиатора в синаптическую щель.
Синаптическая щель – узкая полоска межклеточного пространства, заполненного гелем. В химических синапсах синаптическая щель более широкая, чем в электрических и составляет 10—50 нм.
Важный компонент постсинапса химического синапса – специализированные мембранные белки – рецепторы. Они осуществляют восприятие медиатора и запуск нервного импульса.
Особенности строения химического синапса обусловили закономерности передачи возбуждения:
- односторонность проведения,
- наличие синаптической задержки, связанной с затратой времени на диффузию медиатора,
- утомляемость, вызванная расходом медиатора.
В зависимости от медиатора химические синапсы разделяются на
- катехоламинергические, содержащие биогенные амины (адреналин, норадреналин, дофамин);
- серотонинэргические, содержащие серотонии;
- холинергические, содержащие ацетилхолин;
- ГАМК-ергические, содержащие гамма-аминомасляную кислоту;
- пептидергические, содержащие пептиды и др.
В синапсе не всегда вырабатывается только один медиатор. Обычно основной медиатор выбрасывается вместе с другим, играющим роль модулятора.
По конечному физиологическому эффекту химические синапсы могут быть возбуждающими или тормозными. Если первые способствуют возникновению возбуждения в постсинаптической, то вторые, напротив, прекращают или предотвращают его появление, препятствуют дальнейшему распространению импульса. Обычно тормозными являются глицинергические (медиатор — глицин) и ГАМК-ергические синапсы.
В электрических синапсах отсутствуют синаптические пузырьки, более узкая синаптическая щель, нет специализации пре- и постсинаптической мембраны. Здесь происходит прямая передача электрического импульса с одной клетки на другую. Для облегчения передачи возбуждения клетки соединяются высокопроницаемыми контактами, образованными с помощью особых белковых комплексов - коннексонов (каждый коннексон состоит из шести белковых субъединиц). Расстояние между мембранами клетки в электрическом синапсе 2- 4 нм.
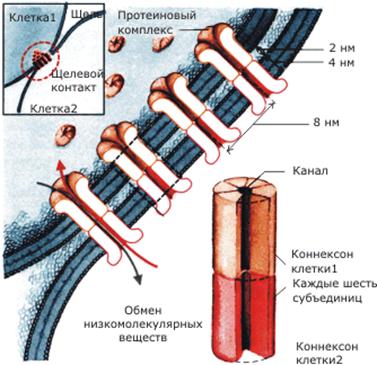
Рис. 2. Строение электрического синапса.
Для нервной системы млекопитающих электрические синапсы менее характерны, чем химические.
Передача возбуждения в электрическом синапсе осуществляется:
- в обе стороны,
- быстрее, чем в электрическом синапсе (без синаптической задержки),
- надежнее.
Электрические синапсы проводят только возбуждающие импульсы, они не утомляемы.
В некоторых синаптических структурах вместе функционируют и электрический и химический механизмы передачи - это так называемые смешанные синапсы.
В соответствии с морфологическим принципом синапсы подразделяют:
§ - аксоаксональные синапсы (между двумя аксонами);
§ - аксодендритические синапсы (между аксоном одного нейрона и дендритом другого);
§ - аксосоматические синапсы (между аксоном одного нейрона и телом другого);
§ - дендродендритические (между дендритами двух или нескольких нейронов);
§ - нервно-мышечные синапсы (между аксоном мотонейрона и мышечным волокном);
§ - нервно-секреторные синапсы (аксо-вазальные - между аксоном и кровеносным сосудом).
Кроме этого, все синапсы делят также на центральные (в головном и спинном мозге) и периферические (нервно-мышечные, аксоэпителиальные и синапсы вегетативных ганглиев).
Нейроэффекторные взаимодействия осуществляются нейромышечными и нейросекреторными окончаниями. Передают нервный импульс с нейрона на ткани рабочего органа. Нервно-мышечный синапс представляет собой контакт многочисленных ветвлений терминали аксона и участка мышечного волокна. Пре– и постсинаптическая мембраны разделены синаптической щелью порядка 50 нм. Пресинаптическое окончание аналогично таковому в межнейронных синапсах. Пузырьки содержат медиатор ацетилхолин. Постсинаптическая мембрана так же, как и в холинэргических межнейронных синапсах, содержит белок холинорецептор. Механизм передачи потенциала действия нервно-мышечного синапса аналогичен таковому в межнейронных синапсах.
Нейросекреторные синапсы устроены значительно проще, чем нервно-мышечные. Они представляют собой лишь утолщение терминалей аксона, содержащее в основном ацетилхолин. Медиатор нервных окончаний поступает непосредственно в межклеточное пространство.
Общий план строения нервной системы человека.
Нервной системой называется совокупность анатомически и функционально взаимосвязанных структур, занимающихся координацией и регуляцией деятельности различных систем и аппаратов организма, обеспечивающих существование организма как единого целого и установление его взаимодействия с внешней средой. Нервную систему человека условно подразделяют по топографическому принципу на центральную и периферическую.

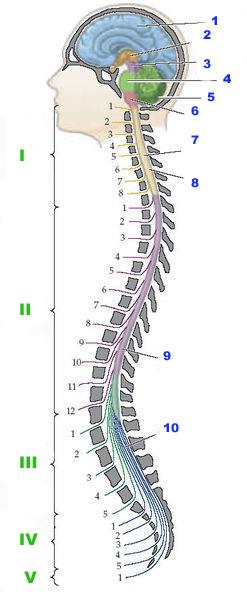
Рис. Общий план строения нервной системы человека:
1 — головной мозг;
2 — спинной мозг;
3 — периферический отдел нервной системы.
К центральной нервной системе (ЦНС) относят спинной и головной мозг, к периферической нервной системе – все нервные структуры, расположенные за пределами ЦНС (корешки, спинномозговые и черепные нервы и их ветви, нервные сплетения и узлы).
Периферическая нервная система связывает ЦНС с рецепторным аппаратом и с эффекторами, передающими нервные импульсы на рабочие органы.
По другой, функциональной, классификации нервную систему подразделяют на соматическую и вегетативную (автономную).
Соматическая нервная система обеспечивает, главным образом, иннервацию тела (сомы), то есть кожи, скелетных мышц. Основная ее функция – восприятие сигналов внешней среды с помощью рецепторов и обеспечение адекватной ответной реакции.
Вегетативная нервная система иннервирует внутренние органы, железы, кровеносные сосуды, а также регулирует обменные процессы в организме. Вегетативная нервная система в свою очередь подразделяется симпатическую и парасимпатическую части. Результаты активности этих двух систем во многом противоположны. Если основная функция симпатической нервной системы состоит в мобилизации организма на борьбу или бегство, то парасимпатическая нервная система преимущественно обеспечивает поддержание гомеостаза.
Таблица. Влияние симпатического и парасимпатического отделов вегетативной нервной системы
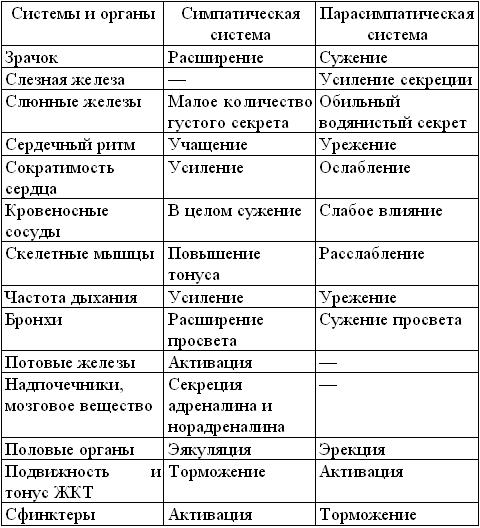
Активация симпатической нервной системы лежит в основе поведения человека, рвущегося в бой. Возбуждение парасимпатической нервной системы обеспечивает пищеварение у человека, лежащего на диване после сытного обеда. Симпатическая нервная система возбуждает, а парасимпатическая тормозит деятельность сердца, первая ослабляет двигательную активность кишечника, вторая - ее усиливает. В то же время они могут действовать и заодно: вместе увеличивают двигательную активность слюнных и желудочных желез, хотя состав секретируемого сока в зависимости от доли участия каждой системы меняется.
Существует теория о генетической детерминированности особенностей вегетативной нервной системы у человека и предрасположенности к некоторым заболеваниям в зависимости от преобладания тонуса одного из ее отделов.
Выделение всех вышеназванных отделов в нервной системе является условным. В действительности нервная система представляет собой анатомически и функционально единое целое.
Общий план строения ЦНС.
Центральная нервная система состоит из спинного и головного мозга. Они надежно защищены от внешних воздействий. Головной мозг заключен в черепную коробку, спинной – позвоночный канал.
В спинном мозге различают шейный, грудной, поясничный, крестцовый и копчиковый отделы.
В головном мозге выделяют следующие отделы:
- продолговатый мозг
- задний (мост и мозжечок)
- средний мозг (четверохолмие и ножки мозга)
промежуточный мозг (таламус, эпиталамус, гипоталамус)
- конечный мозг (базальные ядра и кора).
Рис.1. Общий план строения центральной нервной системы человека.
Головной мозг:
1. конечный мозг
2. промежуточныймозг.
3. средниймозг.
4. Мост.
5. Мозжечок.
6. Продолговатый мозг.
7. Спинной мозг:
I. Шейные нервы.
II. Грудные нервы.
III. Поясничные нервы.
IV. Крестцовые нервы.
V. Копчиковые нервы.
8. Шейное утолщение.
9. Поясничное утолщение.
10. «Конский хвост»
Головной и спинной мозг снаружи защищены тремя мозговыми оболочками, состоящими из соединительной ткани: твердой (dura mater), паутинной (arachnoidea) и мягкой (pia mater). Твердая мозговая оболочка развивается из мезодермы, паутинная и мягкая имеют нейрогенное, эктодермальное происхождение.
Рис.2. Строение оболочек головного мозга (разрез во фронтальной плоскости):
1-твердая оболочка головного мозга;
2-кость черепа;
3-грануляции паутинной оболочки;
4-венозный синус;
5-кожа;
6- вена;
7-паутинная оболочка головного мозга;
8-подпаутинное пространство;
9-мягкая оболочка головного мозга
10-головной мозг;
11-серп большого мозга.
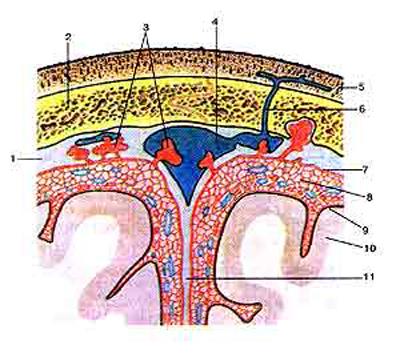
Твердая мозговая оболочка образована плотной соединительной тканью и в головном мозге состоит из двух листков: наружного, прирастающего к костям черепа и внутреннего, который в виде тонкой пластины покрывает поверхность мозга, внедряясь между полушариями и мозжечком, образуя «серпы» твердой мозговой оболочки. В некоторых местах внутренний и наружный листки прилегают друг к другу неплотно, образуя расширения – венозные синусы, заполненные венозной кровью. На наружной поверхности оболочки залегают кровеносные сосуды. В спинном мозге твердая мозговая оболочка отделена от поверхности позвонков узким промежутком – эпидуральным пространством, заполненным жировой тканью. Под твердой мозговой оболочкой находится узкое субдуральное пространство, заполненное спинномозговой жидкостью.
Глубже располагается тонкая паутинная мозговая оболочка, толщина которой в различных участках мозга неодинакова. Она лишена кровеносных сосудов. Паутинная оболочка не заходит в борозды и углубления мозга, перекидываясь через них в виде мостиков. В результате между паутинной и мягкой мозговыми оболочками располагается подпаутинное пространство, которое расширяясь образует местами цистерны. Заполнено подпаутинное пространство спинномозговой жидкостью. В некоторых участках мозга паутинная оболочка имеет особые выросты – грануляции. Они внедряются в полости венозных синусов. Через грануляции спинномозговая жидкость фильтруется в венозное русло.
Мягкая мозговая оболочка тесно прилегает к мозгу, заходя во все борозды и углубления. Она богата кровеносными сосудами. В некоторых местах кровеносные сосуды развиты очень сильно и образуют сосудистые сплетения, которые внедряются в полость мозговых желудочков. Здесь происходит секреция и обмен спинномозговой жидкости.
Спинномозговая жидкость (ликвор) по своему составу резко отличается от других жидкостей организма. Ликвор бесцветная, прозрачная как дистиллированная вода, жидкость, с плотностью 1,006-1,007, со слабощелочной реакцией, рН 7,4 - 7,6. Состоит на 98,9 - 99,0 % из воды и 1,0 - 1,1 % сухого остатка. В нормальном ликворе взрослого человека практически отсутствуют клеточные элементы: в 1 мкл ликвора содержится 0 – 3 клетки. Содержит белок - 0,22 - 0,33 г/л, глюкозу - 2,2 - 4,4 ммоль/л, что примерно вдвое меньше, чем в крови. Спинномозговая жидкостьобразуется эпителиальной выстилкой сосудистых сплетений мягкой мозговой оболочки. Отток спинномозговой жидкости осуществляется в венозную систему через грануляции паутинной мозговой оболочки. Головной мозг содержит в среднем 130-140 мл ликвора. Функции ликвора многообразны. Известно, что функциональное состояние и деятельность головного мозга в значительной мере зависят от состава, физических и биологических свойств спинномозговой жидкости. Защитная функция ликвора проявляется в удалении продуктов распада нервных клеток и токсических веществ. Поддерживая постоянство состава среды, в которой функционируют нейроны мозга, спинномозговая жидкость выполняет гомеостатическую функцию. До настоящего времени не утратила своего значения теория, доказывающая значение ликвора как амортизирующего фактора, выполняющего роль «водяной подушки», обеспечивающей защиту нервных клеток от механических воздействий. Головной и спинной мозг как бы «плавают» в ликворе, и любое механическое воздействие, в частности удары умеренной силы, не вызывают повреждения вещества мозга.
Обмен веществ между кровью и мозгом обеспечивает гематоэнцефалический барьер. Важнейшим компонентом его является эндотелий кровеносных капилляров мозга, который непроницаем для молекул с диаметром более 1,5 нм. Гематоэнцефалическим барьер могут преодолевать только маленькие молекулы, например ионы, глюкоза, незаменимые аминокислоты и жирные кислоты. Благодаря ему большие молекулы, токсины, вирусы и микробы не могут проникнуть в нейрон, что приводит к значительному повышению толерантности (устойчивости) мозга к вирусным инфекциям. В состав гематоэнцефалического барьера входит также базальная мембрана и коллагеновый матрикс мозговых капилляров и глиальная оболочка, образованная астроцитами, окружающими мозговые сосуды. Существует только одна область мозга, где происходит нарушение гематоэнцефалического барьера, — гипоталамус. В нем находятся клетки, секретирующие нейрогормоны. Сосуды непосредственно подходят к секретирующим нейронам, выделяющим свои биологически активные вещества прямо в кровь. Ввиду функциональной необходимости гематоэнцефалический барьер в этом месте нарушается. Гипоталамус можно назвать «ахиллесовой пятой» мозга, поскольку только здесь возможно проникновение инфекций в нервную систему человека.
Наличие гематоэнцефалического барьера при инфекционных поражениях мозговой ткани может препятствовать ее лечению путем введения антибиотиков в кровь. Молекулы лекарства не могут попасть в мозг в нужном количестве и не имеют возможности подойти непосредственно к очагу инфекции. Единственным выходом из этой ситуации остается пункция: лекарство вводится в позвоночный канал в эпидуральное пространство.
Лимфатические сосуды в нервной ткани отсутствуют.
Спинной мозг.
Спинной мозг (medulla spinalis) - наиболее древний отдел нервной системы, в организации которого сохранились черты сегментарного строения. Он имеет вид неравномерной по толщине, сдавленной в передне-заднем направлении, трубки. Заключен в позвоночный канал и окружен оболочками, между которыми циркулирует спинномозговая жидкость. Длина спинного мозга у взрослого человека составляет 43 см. На уровне большого затылочного отверстия он переходит в головной мозг, а заканчивается на уровне 2-3 поясничного позвонка. Масса – от 34 до 38 грамм, что составляет 2% от массы головного мозга. Построен симметрично.
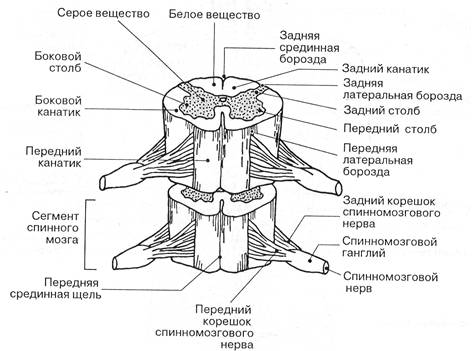
Рис. 1 Строение сегментов спинного мозга
На передней поверхности посередине имеется глубокая передняя срединная щель, на задней поверхности - срединная борозда. По боковой поверхности каждой стороны проходят передняя и задняя латеральные борозды. Они соответствуют местам выхода передних и задних корешков спинномозговых нервов. Передний корешок состоит из отростков двигательных (моторных, эфферентных, центробежных) нервных клеток, расположенных в переднем роге спинного мозга. Задний корешок, чувствительный (афферентный, центростремительный), представлен совокупностью проникающих в спинной мозг центральных отростков чувствительных псевдоуниполярных клеток, тела которых образуют спинномозговой узел (ганглий).
Вблизи спинного мозга в области межпозвоночного отверстия передние и задние корешки соединяются в единый спинномозговой нерв. Каждому сегменту спинного мозга соответствуют четыре корешка спинномозговых нервов или пара соответствующих нервов. Спинной мозг состоит из 31 пары сегментов (8 шейных, 12 грудных, 5 поясничных, 5 крестцовых и 1 копчиковый). Морфологических границ между сегментами спинного мозга не существует, поэтому деление на сегменты является функциональным.
В связи с особенностями развития спинной мозг у взрослых оканчивается на уровне I-II поясничного позвонка, то есть короче позвоночника. Поэтому корешки спинномозговых нервов грудных и особенно пояснично-крестцовых сегментов проходят до соответствующих межпозвоночных отверстий некоторое расстояние внутри позвоночного канала, формируя пучок - конский хвост.
Толщина спинного мозга на всем протяжении неодинакова. В шейном и поясничном отделах имеются шейное и пояснично-крестцовое утолщения. Эти утолщения вызваны увеличением количества нервных элементов, участвующих в иннервации верхних и нижних конечностей. Кроме того спинной мозг имеет изгибы в сагитальной плоскости, соответствующие шейному лордозу и грудному кифозу позвоночного столба.
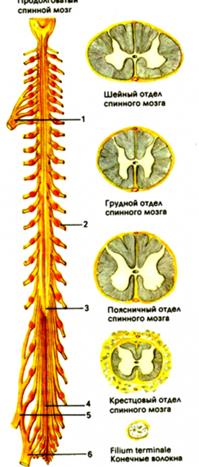
Рис.2. Строение спинного мозга (Справа показано распределение серого и белого вещества в разных отделах спинного мозга.).
Внутреннее строение спинного мозга. Центральное положение в спинном мозге занимает спинномозговой канал. Вокруг него располагается серое вещество. На поперечных срезах оно имеет характерную форму "бабочки". На протяжении спинного мозга форма бабочки несколько меняется (см. рис. 2). Серое вещество представлено преимущественно нервными клетками, которые формируют ядра, сливающиеся в передние, боковые и задние столбы серого вещества. На поперечных срезах выступы серого вещества часто называют рогами. Нейроны в сером веществе располагаются группами. В передних рогах расположены двигательные нейроны, в задних – чувствительные нейроны, а в боковых – нейроны, образующие центры симпатической нервной системы.
По универсальной классификаций серое вещество спинного мозга принято делить на 9 пластин (рис.3), различающихся по структуре и функциональной значимости составляющих их элементов:
I пластина соответствует поверхностно расположенной краевой зоне (zona terminalis) задних столбов спинного мозга. Включает небольшое количество мелких и средних нейронов.
II пластина залегает несколько глубже, здесь расположено губчатое вещество (substantia spongiosa), состоящее из мелких плотно лежащих нейронов.
III пластина соответствует студенистому веществу (substantia gelatinosa). По структуре напоминает предыдущую, содержит нейроны среднего размера. На нейронах II и III пластин оканчивается большое количество афферентов. Назначение этих пластин - конвергенция поступающей информации и передача ее в соответствующие отделы спинного и головного мозга.
IY пластина образована собственными ядрами заднего столба (nucleus proprius cornu posterioris). Их составляют крупные мультиполярные и пучковые нейроны. На них оканчиваются афферентные волокна рецепторов мышц и сухожилий, их аксоны образуют часть спинно-таламических трактов.
Y и YI пластины составляют основание заднего столба и содержат пучковые клетки, образующие грудное ядро (nucleus thoracicus). В этих пластинах формируется основная масса проприоспинальных связей и некоторые восходящие тракты (спинно-мозжечковый тракт).
YII пластина соответствует промежуточному веществу спинного мозга (substantia intermedia). Латеральные отделы на уровне грудных - крестцовых сегментов представлены скоплениями ассоциативных нейронов спинальных центров симпатической и парасимпатической систем и образуют боковые столбы серого вещества спинного мозга.
YIII пластина располагается в медиальной части переднего столба. Отростки нейронов YII и YIII пластин формируют обширные комиссуральные связи, а также восходящие и нисходящие тракты различного типа.
IX пластина включает группу собственных ядер переднего столба спинного мозга (nucleus proprius cornu аnterioris). В них сосредоточены спинальные мотонейроны, управляющие деятельностью скелетной мускулатуры. Группа мотонейронов, связанная с иннервацией определенной мышцы, носит название "мотонейронный пул". Для моторных ядер характерен соматотопический принцип организации.
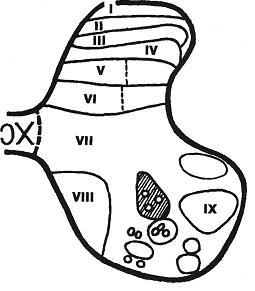
Рис. 3. Пластины серого вещества по классификации Б. Рекседа
Серое вещество спинного мозга образует сегментарный аппарат спинного мозга. Основная его функция – это осуществление врожденных рефлексов в ответ на раздражение (внутреннее или внешнее). Морфологической основой рефлекса является рефлекторная дуга, представленная цепью нейронов, обеспечивающих восприятие раздражения, трансформацию энергии раздражения в нервный импульс, проведение нервного импульса до нервных центров, обработку поступившей информации и peaлизацию ответной реакции. В зависимости от сложности рефлекторного акта различают простые и сложные рефлекторные дуги.
В простой рефлекторной дуге имеются 3 звена: афферентное, вставочное (ассоциативное) и эфферентное.
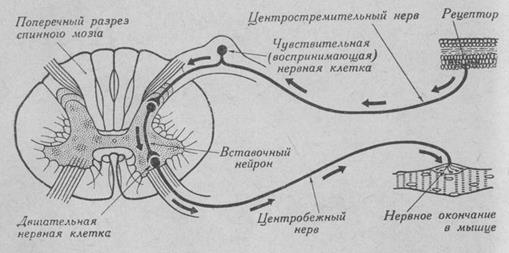
Рис. 4. Схема рефлекторной дуги.
Афферентное звено представлено чувствительным, или рецепторным нейроном, который располагается в спинномозговом узле и является псевдоуниполярной клеткой. От тела этой клетки отходит один отросток. Он вскоре делится на периферический и центральный отростки. Периферический отросток начинается на периферии (в коже, мышцах. сухожилиях, суставных сумках) рецепторами. Область локализации рецепторов, раздражение которой ведет к возникновению определенного рефлекса, называется рефлексогенной зоной. От рефлексогенной зоны возникшие вследствие раздражения нервные импульсы движутся сначала к телу псевдоуниполярной клетки, а затем по ее центральному отростку в спинной мозг. Центральный отросток рецепторного нейрона образует синаптическое окончание на дендритах ассоциативного (вставочного) нейрона. Ассоциативный нейрон представляет собой вставочное звено рефлекторной дуги и является мелкой мультиполярной клеткой с коротким аксоном. Вставочный нейрон получает нервный импульс, своими дендритами или непосредственно поверхностью тела, проводит его по аксону и образует синаптическое окончание на эффекторном нейроне. Эффекторный нейрон (мотонейрон) - это крупная мультиполярная клетка, аксон которой покидает центральную нервную систему и заканчивается эффекторными окончаниями в тканях рабочего органа (в поперечнополосатой мускулатуре).
Усложнение рефлекторных дуг происходит за счет вставочного звена. Ассоциативные нейроны образуют многочисленные ядра (нервные центры) в пределах спинного и головного мозга.
Снаружи серое вещество спинного мозга окружает белое вещество. Оно образовано в основном миелинизированными нервными волокнами. Выделяют передние, боковые и задние канатики белого вещества.
Передний канатик расположен между передней срединной щелью и медиальной поверхностью переднего рога. Боковой - между латеральными поверхностями переднего и заднего рогов. Задний - между срединной бороздой и медиальной поверхностью заднего рога. Каждый канатик состоит из большого количества пучков нервных волокон, которые объединяются в нервные тракты (пути) по общности направления и функционального значения. Нервный тракт - это совокупность нервных волокон, имеющих одно и тоже направление и обеспечивающих передачу одинаковых по функции нервных импульсов.
Проводящие тракты спинного мозга условно подразделяются на проприоспинальные и супраспинальные. Проприоспинальные пути располагаются в пределах спинного мозга, проходят на границе белого и серого вещества и служат для быстрой передачи возбуждения на большое число мотонейронов и интернейронов, связанных с выполнением определенного движения. Ассоциативные проприоспинальные и связывают между собой нейроны одного или разных сегментов спинного мозга одной половины тела. Они начинаются от нейронов серого вещества промежуточной зоны, идут в белое вещество бокового или переднего канатиков спинного мозга и заканчиваются в сером веществе промежуточной зоны или на мотонейронах передних рогов других сегментов. Эти связи выполняют ассоциативную функцию, которая заключается в координации позы, тонуса мышц, движений разных метамеров туловища. К проприоспинальным путям относятся также комиссуральные волокна, соединяющие функционально однородные симметричные и несимметричные участки разных сторон спинного мозга.
Супраспинальные проводящие пути выходят за пределы спинного мозга и обеспечивают его связь со структурами головного мозга. Они разделяются на восходящие (спино-церебральные) и нисходящие (церебро-спинальные).
Восходящие проводящие пути обеспечивают передачу в головной мозг информации от рецепторов тела и внутренних органов. Это сенсорные тракты. Они занимают задний и частично боковой и передний канатики спинного мозга. Задний канатик содержит спинно-бульбарные проводящие пути:
1 - тонкий пучок (fasciculus gracilis) расположен в медиальной части заднего канатика. Он образован аксонами чувствительных нейронов, расположенных в 19 нижних спинномозговых ганглиях и передает информацию от проприорецепторов и частично от тактильных рецепторов нижних конечностей и туловища. Проводятся импульсы сознательной чувствительности. Заканчивается в одноименном ядре продолговатого мозга.
2 - клиновидный пучок (fasciculus cuneatus) находится латеральнее клиновидного. Функционально аналогичен предыдущему. Проводит информацию от рецепторов верхних конечностей и верхней части туловища (от 12 верхних спинномозговых ганглиев).
В боковом канатике располагаются следующие чувствительные тракты:
1 - задний спинно-мозжечковый путь (tractus spinocerebellaris posteroir) располагается в заднелатеральной части бокового канатика, образован аксонами нейронов грудного ядра и проводит импульсы бессознательной проприцептивной чувствительности. Оканчивается на коре мозжечка.
2 - передний спинно-мозжечковый путь (tractus spinocerebellaris anteroir) расположен в переднелатеральной части бокового канатика. Функционально аналогичен предыдущему.
3 - латеральный спинно-таламический путь (tractus spinotalamicus lateralis) расположен медиальнее переднего спинно-мозжечкового тракта. образован аксонами нейронов собственного ядра заднего рога. Функционально связан с проведением болевой и температурной чувствительности. Заканчивается на таламических ядрах промежуточного мозга.
В переднем канатике также находятся несколько восходящих проводящих путей:
1 - передний спинно-таламический путь (tractus spinotalamicus anterior) расположен перед передним столбом серого вещества. Образован нейронами тех же ядер, что и латеральный спинно-таламический путь, проводит импульсы тактильной чувствительности.
2 - спинно-покрышечный путь (tractus spinotectalis) располагается позади передней латеральной борозды спинного мозга. Берет начало от крупных интернейронов II - IY пластин спинного мозга. Заканчивается на ядрах покрышки среднего мозга.
3 - спинно-ретикулярный путь (tractus spinoreticularis) объединяет в своем составе несколько пучков волокон, идущих в ретикулярную формацию ствола мозга. Проводит общую импульсацию от большого количества рецепторов и осуществляют дифференцировку импульсов, проходящих по другим трактам.
Нисходящие проводящие пути связывают головной мозг со спинным. Они осуществляют контроль со стороны высших отделов ЦНС над работой сегментарного аппарата спинного мозга. По локализации источника нисходящие пути делят на пирамидные (кортикоспинальные) и экстрапирамидные тракты. Первые имеют источником кору головного мозга, вторые начинаются в различных областях головного мозга. Нисходящие пути проходят в составе переднего и бокового канатиков спинного мозга.
Передний канатик содержит следующие нисходящие проводящие пути:
1 - передний кортикоспинальный путь (tractus corticospinalis anterior) начинается от пирамидных клеток коры больших полушарий головного мозга. Проходит в переднем отделе переднего канатика. Заканчивается на интернейронах III -IY пластин серого вещества спинного мозга. Посредством их регулируется деятельность мышц, связанная с выполнением позных движений туловища и конечностей. Имеются также многочисленные прямые связи с мотонейронами спинного мозга, что позволяет сознательно контролировать выполнение тонких движений дистальных отделов конечностей.
2 - покрышечно-спинномозговой путь (tractus tectospinalis) расположен в медиальной части переднего канатика, вдоль передней срединной щели. Начинается в области холмов покрышки среднего мозга, заканчивается на мотонейронах собственных ядер передних рогов спинного мозга. Функционально связан с координацией движений головы и туловища в ответ на зрительные и звуковые сигналы. Осуществляет безусловные защитные рефлексы.
3 - ретикуло-спинномозговой путь начинается от аксонов ретикулярной формации моста и продолговатого мозга. Зона окончания обширна (YI-IX пластины). Играет роль в поддержании тонуса мускулатуры.
В боковом канатике проходят следующие нисходящие проводящие пути:
1 - боковой кортико-спинальный путь (tractus corticospinalis lateralis) занимает значительную часть бокового канатика, располагаясь медиодорсально. По происхождению и функционально аналогичен переднему кортикоспинальному пути.
2 - красноядерно-спинальный путь (tractus rubrospinalis) образуется аксонами нейронов красных ядер среднего мозга, проходит в центре передней части бокового канатика. Заканчивается на нейронах собственных ядер передних рогов спинного мозга. Функционально связан с автоматическим, подсознательным управлением движениями и тонусом скелетных мышц.
3 - преддверно-спинномозговой (tractus vestibulospinalis) и оливо-спинномозговой (tractus olivospinalis) пути формируются аксонами нейронов преддверных ядер моста и ядер оливы продолговатого мозга. Проходят в передне-медиальной части бокового канатика и заканчиваются на мотонейронах передних рогов спинного мозга. Функционально связаны с поддержанием равновесия тела и автоматическим изменением положения тела при вестибулярных нагрузках.
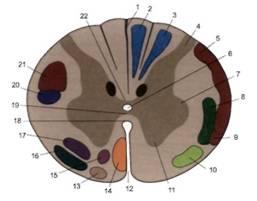
Рис. 5. Проводящие пути на поперечном срезе верхнегрудного отдела спинного мозга.
1 - задняя срединная перегородка;
2 - тонкий пучок;
3 - клиновидный пучок;
4 - задний рог;
5 - задний спиномозжечковый путь,
6 - центральный канал,
7 - боковой рог;
8 -латеральный спиноталамический путь;
9 - передний спиномозжечковый путь;
10 - передний спиноталамический путь;
11 - передний рог;
12 - передняя срединная щель;
13 - оливоспинномозговой путь;
14 - передний корково-спинномозговой (пирамидный) путь;
15 - передний ретикулярно-спинномозговой путь;
16 - преддверно-спинномозговой путь;
17 - ретикулярно-спинномозговой путь;
18 - передняя белая спайка;
19 - серая спайка;
20 - красноядерно-спинномозговой путь;
21 - латеральный корково-спинномозговой (пирамидный) путь;
22 - задняя белая спайка.
Дата добавления: 2015-11-28; просмотров: 3481;