5 страница. Общие закономерности сукцессионного процесса
Общие закономерности сукцессионного процесса. Для любой сукцессии, особенно первичной, характерны следующие общие закономерности протекания процесса.
1. На начальных стадиях видовое разнообразие незначительно, продуктивность и биомасса малы. По мере развития сукцессии эти показатели возрастают.
2. С развитием сукцессионного ряда увеличиваются взаимосвязи между организмами. Особенно возрастает количество и роль симбиотических отношений. Полнее осваивается среда обитания, усложняются цепи и сети питания.
3. Уменьшается количество свободных экологических ниш, и в климаксном сообществе они либо отсутствуют, либо находятся в минимуме. В связи с этим по мере развития сукцессии уменьшается вероятность вспышек численности отдельных видов.
4. Интенсифицируются процессы круговорота веществ, потока энергии и дыхания экосистем.
5. Скорость сукцессионного процесса в большей мере зависит от продолжительности жизни организмов, играющих основную роль в сложении и функционировании экосистем. В этом отношении наиболее продолжительны сукцессии в лесных экосистемах. Короче они в экосистемах, где автотрофное звено представлено травянистыми растениями, и еще быстрее протекают в водных экосистемах.
6. Неизменяемость завершающих (климаксных) стадий сукцессий относительна. Динамические процессы при этом не приостанавливаются, а лишь замедляются. Продолжаются динамические процессы, обусловливаемые изменениями среды обитания, сменой поколений организмов и другими явлениями. Относительно большой удельный вес занимают динамические процессы циклического (флуктуационного) плана.
7. В зрелой стадии климаксного сообщества (не старческой!) биомасса обычно достигает максимальных или близких к максимальным значений. Неоднозначна продуктивность отдельных сообществ на стадии климакса. Обычно считается, что по мере развития сукцессионного процесса продуктивность увеличивается и достигает максимума на промежуточных стадиях, а затем в климаксном сообществе резко уменьшается. Последнее связывают, во-первых, с тем, что в это время максимум первичной продукции потребляется консументами, а во-вторых, экосистема развивает чрезвычайно большую массу ассимиляционного аппарата, что ведет к дефициту освещенности, следствием чего является снижение интенсивности фотосинтеза при одновременном возрастании потерь продуктов ассимиляции на дыхание самих автотрофов.
Эти положения нельзя распространять на все климаксные сообщества. Например, нет реальных предпосылок для увеличения численности гетеротрофов в хвойных лесах (завершающие стадии сукцессий) по сравнению с лиственными (промежуточные стадии). Скорее, в последних больше потребителей зеленой продукции и вероятнее вспышки численности отдельных видов-фитофагов.
Нет также ни теоретических предпосылок,ни фактических данных, которые бы свидетельствовали, что в зрелой климаксной системе, например в еловых лесах, масса хвои достигает чрезмерно (!) высоких значений.
Весь опыт лесоводства свидетельствует о наиболее высокой продуктивности климаксных лесных сообществ (применительно к лесной зоне хвойных или смешанных хвойно-лиственных лесов). В противном случае, с точки зрения получения продукции (древесины), неизбежен вывод о нецелесообразности ориентации на выращивание и сохранение климаксных стадий лесов.
Применительно к другим экосистемам, например луговым, можно согласиться с тем, что возможности получения продукции на климаксной стадии уменьшаются, однако не потому, что сокращается ее нарастание (прирост, продуктивность), а по той причине, что более значительная ее часть отчуждается гетеротрофами в результате образования устойчивых цепей выедания.
Другими словами, продуктивность экосистем на климаксных стадиях сукцессий высока, как правило, максимальна вследствие более полного освоения пространства. Однако возможности снятия человеком первичной продукции лимитируются (иногда до нулевых значений) вследствие включения ее в цепи питания консументов.
IV.8. Стабильность и устойчивость экосистем
Термины «стабильность» и «устойчивость» в экологии обычно рассматриваются как синонимы, и под ними понимается способность экосистем сохранять свою структуру и функциональные свойства при воздействии внешних факторов.
Более целесообразно, однако, разграничивать эти термины, понимая под «стабильностью» данное выше определение, а под «устойчивостью» - способность экосистемы возвращаться в исходное (или близкое к нему) состояние после воздействия факторов, выводящих ее из равновесия. Кроме этого, для более полной характеристики реакции экосистем на внешние факторы целесообразно пользоваться в дополнение к названным еще двумя терминами: «упругость» и «пластичность».
Упругая система способна воспринимать значительные воздействия, не изменяя существенно своей структуры и свойств. Вместе с тем при определенных (запороговых) воздействиях такая система обычно разрушается или переходит в новое качество.
Пластичная система более чувствительна к воздействиям, но она под их влиянием как бы «прогибается» и затем относительно быстро возвращается в исходное или близкое к исходному состояние при прекращении или уменьшении силы воздействия.
Примером упругих экосистем являются климаксные (например, хвойные леса в лесной зоне, коренные тундровые сообщества, типчаково-ковыльные степи и т. п.). Пластичными экосистемами для лесной зоны являются лиственные леса как промежуточные стадии сукцессий. Они, например, выносят в несколько раз больше рекреационных (связанных с посещением населения) и других (пастьба скота, разного рода загрязнения) нагрузок, чем климаксные экосистемы, в которых эдификаторами выступают хвойные виды.
При рассмотрении стабильности и устойчивости как синонимов, обычно считается, что эти качества тем значительнее, чем разнообразнее экосистемы. Данное положение является настолько универсальным, что формулируется как закон: разнообразие - синоним устойчивости (автор Эшби). С этой точки зрения тундровые и пустынные экосистемы рассматриваются как малоустойчивые (нестабильные), а тропические леса, максимально богатые по видовому составу, - как самые устойчивые (стабильные).
Для экосистем с низкой устойчивостью характерны вспышки численности отдельных видов. Последнее связывается с тем, что в маловидовых экосистемах слабо проявляются силы, уравновешивающие численность различных видов (конкуренция, хищничество, паразитизм). Так, для тундровых экосистем типичны периодические резкие увеличения численности мелких грызунов - леммингов. В качестве результата низкой устойчивости этих экосистем рассматривается легкое разрушение их под влиянием внешних воздействий (перевыпаса, технических нагрузок и т. п.). Так, колеи, образующиеся после прохода тяжелой техники (тракторов, вездеходов), сохраняются десятилетиями.
С этих же позиций к неустойчивым и низкостабильным относят агросистемы, создаваемые человеком и представленные обычно одним преобладающим видом растений, интересующим человека. С этой же точки зрения как неустойчивые и нестабильные следует рассматривать сосновые леса на бедных песчаных или щебнистых почвах. Их древесный ярус представлен в таких условиях одним видом (сосной), беден в них и напочвенный (травяной, моховой) покров.
Однако если экосистемы, приведенные выше в качестве примеров, рассматривать с позиций названных выше различии устойчивости и стабильности, то они попадают в разные категории (табл. 4).
Устойчивость, стабильность и другие параметры экосистем зависят часто не столько от структуры самих сообществ (например, их разнообразия), сколько от биолого-экологических свойств видов-эдификаторов и доминантов, слагающих эти сообщества.
Так, высокая стабильность и значительная устойчивость, как видно из табл. 4, присущи сосновым лесам на бедных песчаных почвах, несмотря на малое видовое разнообразие этих экосистем. Это связано, во-первых, с тем, что сосна довольно пластична, и поэтому на изменение условий, например уплотнение почв, она реагирует снижением продуктивности и редко - распадом экосистемы. Однако и в последнем случае, в силу бедности субстрата питательными веществами и влагой, ее молодое поколение не встречает серьезной конкуренции со стороны других видов, и экосистема довольно быстро вновь восстанавливается в том же виде эдафического (почвенного) климакса.
Таблица 4
Характеристики устойчивости и стабильности отдельных экосистем
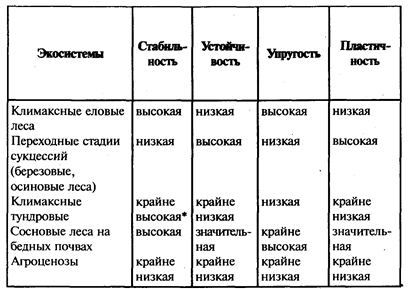
* по основным звеньям: фитоценозам и почвам
Иные параметры устойчивости и стабильности характерны для сосняков на богатых почвах, где они могут сменяться еловыми лесами, обладающими более сильными эдификаторными свойствами. Здесь, несмотря на значительное разнообразие (по видовому составу, ярусности, трофической структуре и т. п.), экосистемы сосновых лесов характеризуются низкой стабильностью и низкой устойчивостью. Сосна в данном случае выступает как промежуточная стадия сукцессионного ряда. Ей удается занимать и удерживать какое-то время такие местообитания только в силу каких-то необычных обстоятельств. Например, после пожаров, когда уничтожаются более сильные конкуренты (ель, лиственные древесные породы).
IV.9. Агроценозы и естественные экосистемы
Основное свойство экосистем - способность естественного развития и прежде всего самовозобновления хотя бы в течение одного-двух поколений. С этой точки зрения нет основания рассматривать агроценозы как экосистемы или одну из стадий (начальную, промежуточную) сукцессионного ряда. Агроценозы сельскохозяйственных культур, особенно однолетних, существуют только при условии постоянного вмешательства человека. При прекращении такого вмешательства обычно начинается вторичная сукцессия с той стадии, которую называют сорняками. Но эта стадия уже не имеет прямого отношения к агроценозу.
Другими словами, такой агроценоз - это совершенно чуждое естественным условиям образование (сообщество), поэтому ему не присуще ни одно из свойств экосистемы (см. табл. 4).
Несколько иные свойства характерны для агроценозов, создаваемых из долгоживущих лесных растений. Эти творения человека можно относить к экосистемам если не на протяжении всей жизни, то на определенных стадиях развития. Здесь наиболее типичны два варианта.
Первый из них относится к созданию леса искусственным путем в условиях, где сукцессии не являются четко выраженными. Например, сосняков на бедных песчаных почвах (см. выше). Здесь вмешательство человека по уходу за выращиваемыми растениями требуется только на начальных этапах жизни, когда сосенки еще настолько малы и слабы, что могут не выдержать конкуренции с травами. В дальнейшем (уже с 3-5 лет жизни) сосна начинает создавать сообщество и постепенно занимать позиции доминантного вида, формирующего свою среду. В последующем образуется сообщество со всеми критериями экосистемы. Правда, некоторые свойства такой экосистемы оказываются не вполне реализованными по сравнению с естественными сообществами. В частности, это проявляется в недостаточной жизнеспособности (устойчивости), обусловленной пониженным, по сравнению с естественными экосистемами, разнообразием. Последнее снижается в результате практически абсолютной выравненности растений по возрасту (возрастная структура как таковая отсутствует) и, в какой-то мере, выравненности почвенного фона в результате обработки почвы, предшествующей посадкам или посевам растений. Пониженная устойчивость проявляется через слабую дифференциацию деревьев по росту и, как следствие этого, ослабление, хотя и в разной степени, всех особей в период смыкания крон и острой внутривидовой конкуренции.
Кроме этого, равномерное размещение растений по площади и создание одновидовых древесных фитоценозов из хвойных видов способствует широко распространенному в настоящее время грибному заболеванию - корневой губке. Последняя имеет следствием очаговую, а порой и полную гибель растительных сообществ как систем.
Второй вариант связан с местопроизрастаниями (прежде всего почвами), характеризующимися значительным богатством питательными веществами и влагой. Здесь создание экосистем, минуя промежуточные стадии сукцессий, требует длительного вмешательства человека в их жизнь. Во всяком случае, до тех пор, пока вводимый вид (например, ель или сосна) не сформирует среду, препятствующую видам-конкурентам (осина, береза, ивы и др.) внедриться в сообщество и захватить главенствующие позиции. В большинстве же случаев победа оказывается на стороне естественных процессов развития экосистем (сукцессий). И виды, вводимые человеком, вытесняются конкурентами полностью или до такой степени, что они не способны сформировать полноценную экосистему с точки зрения целей, которые преследовал человек. Если удается сформировать желаемую экосистему (насаждение), минуя промежуточные стадии сукцессий (обязательно ценой больших затрат), то такие системы, как и в первом случае, оказываются недостаточно устойчивыми.
Исключить эти недостатки искусственных экосистем в значительной мере возможно посредством создания многовидовых сообществ, конечно, при постоянной помощи виду, в котором заинтересован человек. Иногда эти поправки удачно вносит природа через внедрение умеренного количества видов промежуточных стадий сукцессий (береза, осина и др.).
Таким образом, попытки создать сразу климаксные сообщества или их подобие, минуя промежуточные, часто обречены на неудачу по разным причинам. Это должен учитывать человек при решении конкретных хозяйственных проблем. Приведенные выше примеры свидетельствуют, насколько разнообразны взаимосвязи в экосистемах, их зависимость от биотических, абиотических и антропогенных факторов, а также насколько обязателен творческий (системный) подход в каждом конкретном случае, даже при том условии, что выявлены какие-то общие (часто основополагающие) закономерности существования экосистем. Возможности моделирования и создания человеком экосистем во многом зависят не только от биологических свойств видов (в приводимых примерах растительных), но и от условий местопроизрастания. Несомненна также относительность и необходимость конкретизации таких основополагающих экологических постулатов, как «разнообразие - синоним устойчивости», неизбежность резкого снижения продуктивности экосистем в климаксной стадии, неоднозначность терминов «устойчивость» и «стабильность» и др.
Вопросы и задания
1. Из каких основных блоков (звеньев) состоит экосистема? Вспомните определение понятия «Экосистема» из разд.1.
2. Что общего и в чем различаются понятия «Экосистема» и «Биогеоценоз»? Почему каждый биогеоценоз можно назвать экосистемой, но не каждую экосистему можно относить к разряду биогеоценоза, рассматривая последний в соответствии с определением В. Н. Сукачева?
3. Приведите примеры организмов доминантов и эдификаторов. Чем они различаются по роли в экосистемах (биогеоценозах)?
4. Перечислите связи и взаимоотношения между организмами в соответствии с существующими классификациями. Какое значение такие связи имеют для существования экосистем?
5. Что называется «экологической нишей»? Чем это понятие отличается от «местообитания»?
6. Что понимается под трофической структурой экосистем? Что называют трофическим (пищевым) звеном и трофической (пищевой) цепью?
7. Какие энергетические процессы происходят в экосистемах? По каким закономерностям энергия рассеивается и передается в цепях питания? Почему «энергетическая цена» животной пищи существенно выше «энергетической цены» растительной пищи?
8. Что называется продуктивностью и биомассой экосистем? Как связаны эти показатели с воздействием экосистем на среду?
9. Назовите наиболее продуктивные экосистемы суши и океана (пленки жизни и сгущения жизни).
10. Что называется сукцессией? Назовите виды сукцессий. Приведите примеры первичных и вторичных автотрофных и гетеротрофных сукцессий.
11. Как изменяются основные параметры и свойства экосистем в сукцессионном ряду? Перечислите основные закономерности сукцессионного процесса.
12. Чем создаваемые человеком агроценозы отличаются от естественных экосистем (по видовому богатству, устойчивости, стабильности, продуктивности)? Могут ли агроценозы существовать без постоянного вмешательства человека, вложения в них энергии?
V. ПОПУЛЯЦИОННЫЙ УРОВЕНЬ ЖИЗНИ
Если экосистемы - это реально существующие элементы (блоки) биосферы, то популяции в изолированном виде нигде не встречаются в природе. Они входят в состав экосистем и являются одним из элементов их структуры. Популяции выделяются как относительно обособленные части отдельных видов, в пределах которых более вероятны скрещивания и передача информации, чем это возможно между различными популяциями данного вида. Важнейшим фактором обособления популяций внутри вида являются различия условий местообитания. Этот же признак лежит в основе выделения экосистем (биогеоценозов). Поэтому границы популяций часто, хотя далеко не всегда, совпадают с границами экосистем.
В конечном счете экосистему можно рассматривать как сумму популяций (или их частей) разных видов, взаимосвязанных между собой и находящихся в тесном единстве с окружающей средой.
Несмотря на абстрактность вычленения популяций из общих систем (биоценозов, экосистем), им свойственен ряд специфических закономерностей функционирования, важных для существования вида и в целом экосистем различного ранга. На уровне популяций происходят основные адаптации, естественный отбор и эволюционные процессы. Разнообразие популяций внутри вида резко увеличивает его приспособительные способности, освоение среды и, в конечном счете, возможности выживания.
Проявляя заботу о сохранении вида, человек должен прежде всего думать о сохранении популяций. Для популяций различных видов существуют допустимые пределы снижения численности особей, за которыми существование популяции становится невозможным. Точных данных о критических значениях численности популяций в литературе нет. Приводимые значения разноречивы. Остается, однако, несомненным факт, что чем мельче особи, тем выше критические значения их численности. Для микроорганизмов это миллионы особей, для насекомых - десятки и сотни тысяч, а для крупных млекопитающих - несколько десятков. Численность не должна уменьшаться ниже тех пределов, за которыми резко снижается вероятность встречи половых партнеров. Критическая численность также зависит от других факторов. Например, для некоторых организмов специфичен групповой образ жизни (колонии, стаи, стада). Группы внутри популяции относительно обособлены. Могут иметь место такие случаи, когда численность популяции в целом еще достаточно велика, а численность отдельных групп уменьшена ниже критических пределов. Например, колония (группа) перуанского баклана должна иметь численность не менее 10 тыс. особей, а стадо северных оленей - 300-400 голов.
V.1. Структура популяций
Для понимания механизмов функционирования и решения вопросов использования популяций важное значение имеют сведения об их структуре.
Различают половую, возрастную, территориальную и другие виды структуры. В теоретическом и прикладном планах наиболее важны данные о возрастной структуре, под которой понимают соотношение особей (часто объединенных в группы) различных возрастов.
Обычно наибольшей жизнеспособностью отличаются популяции, в которых все возраста представлены относительно равномерно. Такие популяции называют нормальными. Если в популяции преобладают старческие особи, это однозначно свидетельствует о наличии отрицательных факторов в ее существовании, нарушающих воспроизводительные функции. Такие популяции рассматривают как регрессивные, или вымирающие. Требуются срочные меры по выявлению причин такого состояния и их исключению. Популяции, представленные в основном молодыми особями, рассматриваются как внедряющиеся, или инвазионные. Жизненность их обычно не вызывает опасений, но велика вероятность вспышек чрезмерно высокой численности особей, поскольку в таких популяциях не сформировались трофические и другие связи. Особенно опасно, если такие популяции представлены видами, которые здесь ранее отсутствовали. В таком случае популяции обычно находят и занимают свободную экологическую нишу и реализуют свой потенциал размножения, интенсивно увеличивая численность.
Если популяция находится в нормальном или близком к нормальному состоянии, человек из нее может изымать то количество особей или биомассу (последний показатель обычно используется применительно к растительным сообществам), которая прирастает за промежуток времени между изъятиями. Ясно, что изыматься должны прежде всего особи послепродуктивного возраста (окончившие размножение). Если преследуется цель получения определенного продукта, то поправки на возраст, пол или другие характеристики популяций корректируются с учетом поставленной задачи.
Эксплуатация популяций растительных сообществ, в частности, с целью получения древесного сырья обычно приурочивается к тому периоду, когда имеет место возрастное замедление накопления продукции (прироста). Этот период обычно совпадает с максимальным накоплением древесной массы на единице площади.
Количество изымаемой продукции и способ ее изъятия сообразуется с биологическими особенностями популяций. Например, у животных, ведущих групповой образ жизни, как отмечалось выше, нельзя уменьшать численность групп до такой степени, которая повлекла бы за собой потерю ими свойств оптимизации жизненных процессов.
Лесоводами применительно к решаемым задачам и сообразуясь с эколого-биологическими свойствами популяций (экосистем) разработаны различные виды рубок. Прежде всего, они делятся на две большие группы: промежуточного и главного пользования. Рубки промежуточного пользования проводятся практически во всех возрастах жизни леса. При их проведении, наряду с изъятием продукции, преследуется цель создания более благоприятных условий для жизнедеятельности и роста остающейся части древостоя. Ими же создаются условия для увеличения доли более ценных с точки зрения целей хозяйства видов (например, хвойных в смешанных хвойно-лиственных сообществах).
При рубках главного пользования убирается весь древостой, достигший возраста спелости. Этот урожай может сниматься единовременно (сплошные рубки) или в несколько приемов (постепенные, выборочные рубки). Изымается при этом такая часть древостоя, которая не нарушила бы жизнедеятельности популяций и экосистем в целом, механизмов их самоподдержания и соморегулирования (гомеостаза). Такой тип ведения хозяйства рассматривают как мягкое управление природными процессами.
С целью создания условий для появления молодого поколения леса при сплошных рубках применяются такие лесоводственные приемы, как вырубка последовательно небольшими площадями (лесосеками). В таком случае остающиеся рядом с вырубкой древостой являются источником семян, умеренного притенения появляющегося молодого поколения леса, препятствием буйному росту конкурентов из трав, кустарников и нежелательных древесных растений. Появлению молодого поколения леса способствует также оставление на вырубках отдельных, как правило, лучших деревьев, которые выполняют роль обсеменителей и носят название семенников.
Однако в обширных лесных массивах Севера и других регионов часто проводятся так называемые концентрированные рубки большими площадями без учета возможностей восстановления их молодым поколением леса. Они проводятся с использованием тяжелой техники, сопровождаются сильным разрушением и уплотнением почвенного покрова. Это, в свою очередь, ведет, как правило, к цепным реакциям природных процессов, в частности, сложившиеся круговороты воды сменяются накоплением застойных вод на поверхности почв с последующей сменой лесных экосистем болотными. В других случаях, например на песчаных почвах, следствием подобного вмешательства в экосистемы является опустынивание или полное разрушение экосистемы. Такой тип ведения хозяйства рассматривается как жесткое вмешательство в природные процессы. Оно не должно иметь места в деятельности человека.
V.2. Динамика популяций. Гомеостаз
К числу важнейших свойств популяций относится динамика свойственной им численности особей и механизмы ее регулирования. Всякое значительное отклонение численности особей в популяциях от оптимальной связано с отрицательными последствиями для ее существования. В связи с этим популяции обычно имеют адаптационные механизмы, способствующие как снижению численности, если она значительно превышает оптимальную, так и ее восстановлению, если она уменьшается ниже оптимальных значений.
Каждой популяции и виду в целом свойственен так называемый биотический потенциал, под которым понимают теоретически возможное потомство от одной пары особей при реализации способности организмов к биологически обусловленному размножению. Обычно биотический потенциал тем выше, чем ниже уровень организации организмов. Так, дрожжевые клетки, размножающиеся простым делением, при наличии условий для реализации биотического потенциала могли бы освоить все пространство земного шара за несколько часов; гриб дождевик, приносящий до 7,5 млрд. спор, уже во втором поколении освоил бы весь земной шар. Крупным организмам с низким потенциалом размножения потребовалось бы для этого несколько десятилетий или столетий.
Однако биотический потенциал реализуется организмами со значительной степенью полноты только в отдельных случаях и в течение коротких промежутков времени. Например, если быстроразмножающиеся организмы (насекомые, микроорганизмы) осваивают какой-либо субстрат или среду, где нет конкурентов. Такие условия создаются, в частности, при освоении экскрементов крупных животных насекомыми, при размножении организмов в средах, богатых питательными веществами, например в загрязненных органическими или биогенными веществами водоемах и т. п. В этом случае увеличение численности идет по j-образной кривой. Такой тип роста носит название экспоненциального. Близкий к экспоненциальному тип роста характерен в настоящее время для популяции человека. Он обусловлен прежде всего резким снижением смертности в детском возрасте. Для человека характерна кривая увеличения численности первого типа (рис. 6).
Рис. 6
Экспоненциальная (А) и логистическая (Б) кривые роста популяций
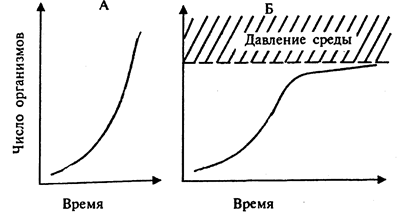
Для большинства же популяций и видов рост численности характеризуется кривой второго типа, которая отражает высокую смертность молодых особей или их зачатков (яйца, икринки, споры, семена и т. п.). При таком типе выживаемости (смертности) численность популяции обычно выражается s-образнои кривой (см. рис.6). Такая кривая носит название логистической. Но и в этом случае периодические колебания численности особей значительны. Такие отклонения от средней численности имеют сезонный (как у многих насекомых), взрывной (как у некоторых грызунов - лемминги, белки) или сглаженный (как у крупных млекопитающих) характер. Численность при этом может существенно отличаться от средних значений: для насекомых - в 107- 106 раз, для позвоночных животных, например грызунов, - в 105 - 106 раз.
Периоды резкого изменения численности носят название «популяционных волн», «волн жизни», «волн численности». Причины таких колебаний до конца не ясны. В одних случаях их связывают с пищевым фактором, в других - с погодными (климатическими) явлениями (например, для леммингов - с количеством тепла, приносимого Гольфстримом), в третьих - с солнечной активностью или комплексом взаимосвязанных факторов, что наиболее вероятно.
Резкие изменения численности относительно средних значений имеют обычно отрицательные следствия для жизни популяций: при высокой численности - из-за ослабления всех особей в результате недостатка пищи, самоотравления среды, возможных массовых заболеваний и т. п.; при низкой численности - из-за превышения порога ее минимальных значений.
В целом можно выделить динамику популяций, независимую от плотности (численности) ее особей и зависимую от плотности. Для первого типа характерна отмеченная выше экспоненциальная кривая роста. Для второго - логистическая кривая. Соответственно различаются и механизмы (факторы), оказывающие влияние на плотность (численность) особей. При независимом от плотности типе изменения численности последняя обусловливается в основном абиотическими факторами (погодные явления, наличие пищи, различного рода катастрофы и т. п.). Эти факторы могут обеспечивать условия как для неограниченного, хотя и кратковременного роста популяций, так и для снижения их численности до нулевой. Такие факторы обычно называют модифицирующими (лат. модификацио-изменение).
Зависимая от плотности динамика популяций обеспечивается биотическими факторами. Их называют регулирующими. Они «работают» по принципу обратной отрицательной связи: чем значительнее численность. Тем сильнее срабатывают механизмы, обусловливающие ее снижение, и наоборот - при низкой численности сила этих механизмов ослабевает и создаются условия для более полной реализации биотического потенциала. Факторы такого типа лежат в основе популяционного гомеостаза, обеспечивающего поддержание численности в определенных границах значений.
Дата добавления: 2016-01-03; просмотров: 2292;