3 страница. 11. Что является центральным звеном биосферы?
11. Что является центральным звеном биосферы?
IV. ЭКОСИСТЕМНЫЙ УРОВЕНЬ ЖИЗНИ
Экосистемы, как отмечалось в разделе («Основные понятия...»), являются основными звеньями (блоками) биосферы. Там же дается определение термина. На уровне экосистем, особенно элементарном (неделимом), представляется возможным рассмотреть более детально, глубоко и последовательно, чем это сделано на примере биосферы, основные свойства и закономерности их функционирования, важные как в теоретическом плане, так и для решения прикладных задач.
Экосистемы в конечном счете являются основными объектами (ячейками) научно обоснованного природопользования, особенно возобновимыми ресурсами. Их уровень используется для обоснования допустимых объемов изъятия продукции (при сохранении устойчивости), увеличения ее выхода (повышения продуктивности) и для решения других вопросов. В конечномсчете через сохранение отдельных элементарных экосистем решается важнейшая проблема современности - предотвращения или нейтрализации неблагоприятных явлений глобального кризиса, сохранения биосферы в целом.
IV. 1. Организация (структура) экосистем
Для того чтобы экосистемы функционировали (существовали) неограниченно долго и как единое целое, они должны обладать свойствами связывания и высвобождения энергии, а также круговоротом веществ. Экосистема, кроме этого, должна иметь механизмы, позволяющие противостоять внешним воздействиям (возмущениям, помехам), гасить их. Для раскрытия этих механизмов познакомимся с различными видами структур и другими характеристиками (свойствами) экосистем.
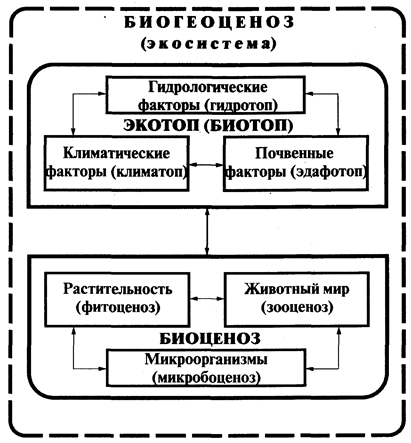
Блоковая модель экосистемы. Любая экосистема состоит из двух блоков. Один из них представлен комплексом взаимосвязанных живых организмов - биоценозом, а второй - факторами среды - биотопом или экотопом. В таком случае можно записать: экосистема = биоценоз + биотоп (экотоп). В. Н. Сукачев блоковую модель в ранге биогеоценоза в виде схемы изобразил на рис. 2.
Этот рисунок позволяет наглядно представить, чем отличаются понятия «экосистема» и «биогеоценоз», на что мы обращали внимание в разделе «Основные понятия...». Биогеоценоз, по В. Н. Сукачеву, включает все названные блоки и звенья. Это понятие обычно используют применительно к сухопутным системам. В биогеоценозах обязательно наличие в качестве основного звена растительного сообщества (фитоценоза). Примеры биогеоценозов - однородные участки леса, луга, степи, болота и т. п.
Экосистемы могут и не иметь растительное звено. Таким примером являются системы, формирующиеся на базе разлагающихся органических остатков, гниющих в лесу деревьев, трупов животных и т. п. В них достаточно присутствие зооценоза и микробоценоза или только микробоценоза, способных осуществлять круговорот веществ.
Таким образом, каждый биогеоценоз может быть назван экосистемой, но не каждая экосистема относится к рангу биогеоценоза.
Чтобы снять терминологические неясности, соавтор В. Н. Сукачева по формированию науки биогеоценологии - профессор В. Н. Дылис - образно определил биогеоценоз как экосистему, но только в рамках фитоценоза.
Биогеоценозы и экосистемы могут различаться и по временному фактору (продолжительности существования). Любой биогеоценоз потенциально бессмертен, поскольку все время пополняется энергией за счет деятельности растительных фото- или хемосинтезирующих организмов. В то же время экосистемы без растительного звена заканчивают свое существование одновременно с высвобождением в процессе разложения субстрата всей содержащейся в нем энергии. Надо, однако, иметь в виду, что в настоящее время термины «экосистема» и «биогеоценоз» нередко рассматриваются как синонимы.
Видовая структура экосистем. Под видовой структурой понимается количество видов, образующих экосистему, и соотношение их численностей. Точных данных о количестве видов в экосистемах нет. Это связано с тем, что трудно учесть видовое разнообразие всех мелких организмов (особенно микроорганизмов). Оно исчисляется сотнями и десятками сотен. Видовое разнообразие обычно тем значительнее, чем богаче условия (биотоп) экосистемы. В этом отношении самыми богатыми по видовому разнообразию являются, например, экосистемы дождевых тропических лесов. Только древесные виды исчисляются в них сотнями.
Рис.2
Схема биогеоценоза (экосистемы), по В.Н.Сукачеву
Богатство видов зависит также от возраста экосистем. Молодые экосистемы, возникающие, например, на таком изначально безжизненном субстрате, как отвалы пород, извлекаемые из глубинных слоев земной коры при добыче полезных ископаемых, крайне бедны видами. В дальнейшем по мере развития экосистем их видовое богатство увеличивается. Но в хорошо сформировавшихся экосистемах оно может несколько уменьшаться. К тому времени обычно выделяется один или 2-3 вида, которые явно преобладают по численности особей. Например, в еловом лесу - ель, в смешанном - ель, береза и осина, в степи - ковыль и типчак. Эти виды занимают большую часть пространства, оставляя меньше места для других видов.
Виды, явно преобладающие по численности особей, носят название доминантных (лат. доминантис - господствующий). Наряду с доминантами в экосистемах выделяются виды-эдификаторы (лат. эдификатор - строитель). К ним относят те виды, которые являются основными образователями среды. Обычно вид-доминант одновременно является и эдификатором. Например, ель в еловом лесу наряду с доминантностью обладает высокими эдификаторными свойствами. Они выражаются в ее способности сильно затенять почву, создавать кислую среду своими корневыми выделениями и при разложении мертвого органического вещества, образовывать специфические для кислой среды подзолистые почвы. Вследствие высоких эдификаторных свойств ели под ее пологом могут жить только виды растений, которые способны мириться со скудным освещением (теневыносливые и тенелюбивые). В то же время под пологом елового леса доминантным видом может быть, например, черника, но она не является существенным эдификатором.
Видовое разнообразие - очень важное свойство экосистем. С ним, как отмечалось выше, связана устойчивость систем к неблагоприятным факторам среды. Разнообразие обеспечивает как бы подстраховку, дублирование устойчивости. Вид, который присутствует в числе единичных экземпляров, при неблагоприятных условиях для широко представленного вида, в том числе и доминантного, может резко увеличить свою численность и таким образом заполнить освободившееся пространство (экологическую нишу), сохранив экосистему как единое целое.
Видовую структуру обычно используют для оценки условий местопроизрастания по растениям-индикаторам. Так, для лесной зоны кислица указывает на условия увлажнения, близкие к оптимальным, и значительное богатство почв питательными минеральными веществами; черника - на несколько избыточное увлажнение и некоторый дефицит элементов минерального питания; брусника - на дефицит увлажнения и почвенного плодородия; мхи (кукушкин лен и особенно сфагнум) - на чрезмерно избыточное увлажнение, дефицит минеральных веществ, недостаток кислорода для дыхания корней и наличие процессов торфообразования. Наряду с индикаторами меняется состав и других видов, произрастающих под пологом эдификаторов.
Названия экосистем (биогеоценозов). По растениям-эдификаторам или доминантам и растениям-индикаторам обычно называют биогеоценозы (экосистемы). Лесоводы их определяют как типы леса (например, ельники-кисличники, ельники-черничники, ельнико-сфагновые и др.). По такому же принципу классифицируются и называются другие экосистемы. Например, для степей выделяются типчаково-ковыльные, злаково-разнотравные и другие системы.
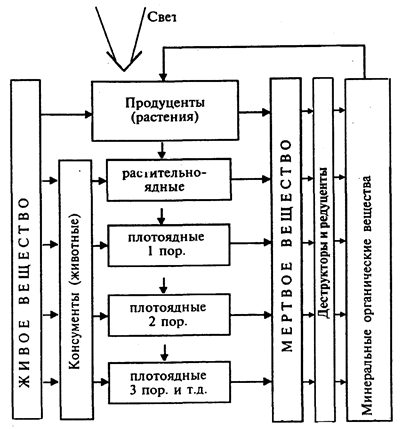
Трофическая (функциональная) структура экосистем. Цепи питания. Любая экосистема включает несколько трофических (пищевых) уровней или звеньев. Первый уровень представлен растениями. Их называют автотрофами (греч. аутос - сам; трофо - пища) или продуцентами (лат. продуцена - создающий). Второй и последующие уровни представлены животными. Их называют гетеротрофами (греч. геторос - другой) или консумен-тами. Последний уровень в основном представлен микроорганизмами и грибами, питающимися мертвым веществом. Их называют редуцентами (лат. редуцере - возвращать). Они разлагают органическое вещество до исходных минеральных элементов.
Взаимосвязанный ряд трофических уровней представляет цепь питания, или трофическую цепь (рис. 3). Главное свойство цепи питания - осуществление биологического круговорота веществ и высвобождение запасенной в органическом веществе энергии. Важно подчеркнуть, что цепь питания не всегда может быть полной. В ней могут отсутствовать растения (продуценты). Такая цепь питания характерна, как отмечалось выше, для сообществ, формирующихся на базе разложения животных или растительных остатков, например, накапливающихся в лесах на почве (лесная подстилка).
В цепи питания очень часто отсутствуют или представлены небольшим количеством животные (гетеротрофы). Например, в лесах отмирающие растения или их части (ветви, листья и др.) сразу включаются в звено редуцентов, которые завершают круговорот.
Исходя из положения: разнообразие - синоним устойчивости, можно заключить, что экосистемы с более длинными цепями питания характеризуются повышенной надежностью и более интенсивным круговоротом веществ.
IV.2. Связи организмов в экосистемах
Ни один организм в природе не существует вне связей со средой и другими организмами. Эти связи - основное условие функционирования экосистем. Через них, как было показано выше, осуществляется образование цепей питания, регулирование численности организмов и их популяций, реализация механизмов устойчивости систем и другие явления. В процессе взаимосвязей происходит поглощение и рассеивание энергии и в конечном счете осуществляются средообразующие, средоохранные и средостабилизирующие функции систем.
Подобные экосистемные связи обусловлены всем ходом эволюционного процесса. По этой причине и любое их нарушение не остается бесследным, требует длительного времени для восстановления. В связи с этим экологически обусловленное поведение человека в природе невозможно без знакомства с этими связями и последствиями их нарушения. Целесообразно выделять взаимосвязи и взаимоотношения организмов в природе (экосистемах) как различные понятия.
Взаимосвязи организмов. Взаимосвязи обычно классифицируются по «интересам», на базе которых организмы строят свои отношения.
Самый распространенный тип связей базируется на интересах питания. Такие связи носят название пищевых или трофических (греч. трофо - питание). В данный тип связей выделяется питание одного организма другим или продуктами его жизнедеятельности (например, экскрементами), питание сходной пищей (например, мертвым органическим веществом). Этим типом связей объединяются растения и насекомые, опыляющие их цветки. На базе трофических связей возникают цепи питания.
Связи, основанные на использовании местообитаний, носят название топических (греч. топос - место). Например, топические связи возникают между животными и растениями, которые предоставляют им убежище или местообитание (насекомые, прячущиеся в расщелинах коры деревьев или живущие в гнездах птиц, растения, поселяющиеся на стволах деревьев (но не паразиты). Не только трофическими, но и топическими отношениями связаны паразиты с организмами, на которых они паразитируют.
Рис.3
Трофическая (функциональная) структура экосистемы (цепь питания) и круговорот вещества в ней
Следующий тип связей носит название форических (лат. форас -наружу, вон). Они возникают в том случае, если одни организмы участвуют в распространении других или их зачатков (семян, плодов, спор). Животными это распространение может осуществляться как на наружных покровах, так и в пищеварительном тракте.
Выделяют также тип связей, которые носят название фабрических (лат. фабрикатио - изготовление). Для них характерно использование одними организмами других или продуктов их жизнедеятельности, частей (например, растений, перьевого покрова, шерсти, пуха) для постройки гнезд, убежищ и т. п.
Взаимоотношения организмов. Данная классификация строится по принципу влияния, которое оказывают одни организмы на другие в процессе взаимных контактов. Эти взаимоотношения можно обозначить математическими значками «+», «–», «0» (положительно, отрицательно, нейтрально).
Если взаимоотношения обоим партнерам выгодны, они обозначаются значками (+,+) и носят название симбиоза или мутуализма. Степень этих связей различна. В ряде случаев организмы настолько тесно связаны, что функционируют как единый организм. Например, лишайники, представляющие симбиоз гриба и водоросли. Водоросль поставляет грибу продукты фотосинтеза, а гриб для водоросли является поставщиком минеральных веществ и, кроме того, субстратом, на котором она живет. В то же время сожительство грибов с корнями растений (микориза) носит хотя и взаимовыгодные, но не в такой степени тесные взаимоотношения. Тип взаимовыгодных отношений широко распространен. Сюда относятся и микроорганизмы, населяющие пищеварительный тракт животных, способствуя усвоению пищи; и, в ряде случаев, травоядные животные. Установлено, что исключение поедания трав животными может иметь следствием оскудение растительных сообществ, снижение ими продуктивности и устойчивости. Даже умеренное объедание листьев древесных растений насекомыми или их гусеницами может быть положительным не только для животных, но и для растений.
Взаимоотношения, которые положительны для одного вида и отрицательны для другого (+,-), характеризуются как хищничество и паразитизм. Хищник и паразит обычно приспосабливаются к использованию других организмов (их жертв и хозяев), а последние, в свою очередь, имеют адаптации, которые сохраняют им жизнь. Эти типы взаимоотношений обычно играют большую роль в регулировании численности организмов. Интенсивное размножение хищников и паразитов обычно имеет следствием уменьшение численности их жертв или хозяев.
В свою очередь, уменьшение численности жертв и хозяев подрывает кормовую базу хищников и паразитов, что ведет к сокращению их численности и т. д. В конечном счете имеет место обычно пульсирующая численность организмов, вступающих в такие типы взаимоотношений.
Хотя взаимоотношения типа хищничества и паразитизма сходны по результатам влияния на численность особей, они резко различаются по образу жизни и адаптациям. Во взаимоотношениях хищник-жертва оба организма постоянно совершенствуются: первый в плане успешности охоты, второй - в отношении самосохранения. И в том и в другом случае требуется быстрая реакция, высокая скорость передвижения, хорошее зрение, обоняние и т. п.
Во втором типе взаимоотношений у паразита адаптации идут по пути специализации структур на использование хозяина как источника пищи и «благоустроенного» местообитания. Результатом этого является упрощение многих органов (пищеварительный тракт, накожные покровы, органы передвижения, чувств и др.). Вместе с тем, поскольку жизнь паразита очень тесно связана с хозяином, он адаптирован на выживание во внешней среде после смерти хозяина. Достигается это за счет большого количества зачатков (семян, спор, цист и т. п.), обычно долго сохраняющихся в среде.
Адаптации хозяина направлены, как правило, на уменьшение вреда от паразита. Это проявляется в выработке активного иммунитета, заключении внутренних паразитов в различного вида капсулы (галлы, цецидии и т. п.).
В ряде случаев адаптации паразитов и хозяев приводят к их взаимовыгодным отношениям типа симбиоза. Есть основание полагать, что в большинстве случаев симбиоз (мутуализм) вырос из паразитизма.
Взаимоотношения, невыгодные обоим партнерам (-,-), носят название конкуренции. Последняя тем сильнее, чем ближе потребности организмов к фактору или условию, за которые они конкурируют. В этом отношении наиболее близки интересы организмов одного вида, и, следовательно, внутривидовая конкуренция рассматривается как более острая по сравнению с межвидовой. Однако данное положение противоречит тому факту, что практически все механизмы существования вида направлены на его выживание. Такое противоречие решается тем, что на внутривидовом уровне есть механизмы, которые позволяют снять остроту конкурентной борьбы, в том числе жертвуя частью особей (см. разд.V.2). Конкуренция и взаимоотношения типа хищник-жертва являются основными в совершенствовании видов, в то время как взаимоотношения типа мутуализма (симбиоза) способствуют оптимизации жизненных процессов, более полному освоению среды.
Менее распространенным типом взаимоотношений является комменсализм (франц. комменсал - сотрапезник) - отношения, положительные для одного и безразличные для другого партнера (+,0), его иногда делят на нахлебничество, когда один организм поедает остатки пищи со «стола» другого (крупного) организма (например, акулы и сопровождающие их мелкие рыбы; львы и гиены) и квартиранство, или синойкийю (греч. синойкос -сожительство), когда одни организмы используют другие как «квартиру», убежище. Например, молодь некоторых морских рыб прячется под зонтик из щупалец медуз, или некоторые насекомые живут в норах животных, гнездах птиц, используя их только для укрытия.
Не часто встречается также аменсализм (лат. аменс - безрассудный, безумный) - отрицательный для одного организма и безразличный для другого (-,0). Например, светолюбивое растение, попавшее под полог леса. Отношения, при которых организмы, занимая сходные местообитания, практически не оказывают влияния друг на друга, носят название нейтрализма (0,0). Например, белки и лоси в лесу. Сохранение разнообразия связей - важнейшее условие устойчивости экосистем.
IV.3. Экологическая ниша
Для понимания различного вида существующих связей в экосистемах и обусловленности механизмов их функционирования важно познакомиться с одним из основополагающих понятий экологии - экологической нишей.
Каждый вид или его части (популяции, группировки различного ранга) занимают определенное место в окружающей их среде. Например, определенньш вид животного не может произвольно менять пищевой рацион или время питания, место размножения, убежища и т. п. Для растений подобная обусловленность условий выражается, например, через светолюбие или тенелюбие, место в вертикальном расчленении сообщества (приуроченность к определенному ярусу), время наиболее активной вегетации. Например, под пологом леса одни растения успевают закончить основной жизненный цикл, завершающийся созреванием семян, до распускания листьев древесного полога (весенние эфемеры). В более позднее время их место занимают другие, более теневыносливые растения. Особая группа растений способна на быстрый захват свободного пространства (растения-пионеры), но отличается низкой конкурентной способностью и поэтому быстро уступает свое место другим (более конкурентоспособным) видам.
Приведенные примеры иллюстрируют экологическую нишу или отдельные ее элементы. Под экологической нишей понимают обычно место организма в природе и весь образ его жизнедеятельности, или, как говорят, жизненный статус, включающий отношение к факторам среды,видам пищи, времени и способам питания, местам размножения, укрытий и т. п. Это понятие значительно объемнее и содержательнее понятия «местообитание». Американский эколог Одум образно назвал местообитание «адресом» организма (вида), а экологическую нишу - его «профессией». На одном местообитании живет, как правило, большое количество организмов разных видов. Например, смешанный лес - это местообитание для сотен видов растений и животных, но у каждого из них своя и только одна «профессия» - экологическая ниша. Так, сходное местообитание, как отмечалось выше, в лесу занимают лось и белка. Но ниши их совершенно разные: белка живет в основном в кронах деревьев, питается семенами и плодами, там же размножается и т. п. Весь жизненный цикл лося связан с подпологовым пространством: питание зелеными растениями или их частями, размножение и укрытие в зарослях и т. п.
Если организмы занимают разные экологические ниши, они не вступают обычно в конкурентные отношения, сферы их деятельности и влияния разделены. В таком случае отношения рассматриваются как нейтральные.
Вместе с тем в каждой экосистеме имеются виды, которые претендуют на одну и ту же нишу или ее элементы (пищу, укрытия и пр.). В таком случае неизбежна конкуренция, борьба за обладание нишей. Эволюционно взаимоотношения сложились так, что виды со сходными требованиями к среде не могут длительно существовать совместно. Эта закономерность не без исключений, но она настолько объективна, что сформулирована в виде положения, которое получило название «правило конкурентного исключения». Автор этого правила эколог Г. Ф. Гаузе. Звучит оно так: если два вида со сходными требованиями к среде (питанию, поведению, местам размножения и т. п.) вступают в конкурентные отношения, то один из них должен погибнуть либо изменить свой образ жизни и занять новую экологическую нишу. Иногда, например, чтобы снять острые конкурентные отношения, одному организму (животному) достаточно изменить время питания, не меняя самого вида пищи (если конкуренция возникает на почке пищевых отношений), или найти новое местообитание (если конкуренция имеет место на почве данного фактора) и т. п.
Из других свойств экологических ниш отметим, что организм (вид) может их менять на протяжении своего жизненного цикла. Наиболее яркий пример в этом отношении - насекомые. Так, экологическая ниша личинок майского жука связана с почвой, питанием корневыми системами растений. В то же время экологическая ниша жуков связана с наземной средой, питанием зелеными частями растений.
Сообщества (биоценозы, экосистемы) формируются по принципу заполнения экологических ниш. В природном сформировавшемся сообществе обычно все ниши заняты. Именно в такие сообщества, например в долгосуществующие (коренные) леса, вероятность внедрения новых видов очень мала. В то же время следует иметь в виду, что занятость экологических ниш в определенной мере понятие относительное. Все ниши обычно освоены теми организмами, которые характерны для данного региона. Но если организм приходит извне (например, заносятся семена или другие зачатки) случайно или преднамеренно, например в результате внедрения человеком новых видов (интродукция, акклиматизация), то он может найти для себя свободную нишу в связи с тем, что на нее не было претендентов из набора существующих видов. В таком случае обычно неизбежно быстрое увеличение численности (вспышка) вида-пришельца, поскольку он находит крайне благоприятные условия (свободную нишу) и, в частности, не имеет врагов (хищников, паразитов или других организмов, которые им питаются). Такие явления не единичны. Например, размножение кроликов, завезенных в Австралию; перемещение ондатры из Азии в европейскую часть; интенсивное продвижение колорадского жука в новые районы.
С экологическими нишами в значительной мере связаны жизненные формы организмов. К последним относят группы видов, часто систематически далеко отстоящие, но выработавшие одинаковые морфологические адаптации в результате существования в сходных условиях. Например, сходством жизненных форм характеризуются дельфины (млекопитающие) и интенсивно передвигающиеся в водной среде хищные рыбы. В условиях степей сходными жизненными формами представлены тушканчики и кенгуру (прыгуны). В растительном мире отдельными жизненными формами представлены многочисленные виды деревьев, занимающие в качестве нити верхний ярус, кустарники, существующие под пологом леса, и травы - в напочвенном покрове.
IV.4. Энергетика экосистем
Живые организмы, входящие в экосистемы, для своего существования должны постоянно пополнять и расходовать энергию. Растения, как известно, способны запасать энергию в химических связях в процессе фотосинтеза или хемосинтеза. При фотосинтезе связывается только энергия с определенными длинами волн -380-710 нм. Эту энергию называют фотосинтетически активной радиацией (ФАР). Она по длинам волн близка к видимой части спектра. На эту радиацию обычно приходится около 40% общей солнечной радиации, достигающей земной поверхности. Остальная часть спектра относится либо к более короткой (ультрафиолетовой), либо к более длинной (инфракрасной) радиации. С последней обычно связан тепловой эффект.
Растения в процессе фотосинтеза связывают лишь небольшую часть солнечной радиации. Даже по отношению к фотосинтетически активной - это в среднем для земного шара менее 1%. Только наиболее продуктивные экосистемы, такие как плантации сахарного тростника, тропические леса, посевы кукурузы, в оптимальных условиях могут связывать до 3-5% ФАР. В опытах с кондиционированными условиями по всем факторам среды за короткие периоды времени удавалось достичь эффективности фотосинтеза по усвоению солнечной энергии порядка 8-10% ФАР.
Растения являются первичными поставщиками энергии для всех других организмов в цепях питания. Существуют определенные закономерности перехода энергии с одного трофического уровня на другой вместе с потребляемой пищей. Основная часть энергии, усвоенной консументом с пищей, расходуется на его жизнеобеспечение (движение, поддержание температуры тела и т. п.). Эту часть энергии рассматривают как траты на дыхание, с которым в конечном счете связаны все возможности ее высвобождения из химических связей органического вещества.
Часть энергии переходит в тело организма-потребителя вместе с увеличивающейся массой (приростом, продукцией). Некоторая доля пищи, а вместе с ней и энергия не усваиваются организмом. Они выводятся в окружающую среду вместе с продуктами жизнедеятельности (экскрементами). В последующем эта энергия высвобождается другими организмами, которые потребляют продукты выделения.
Баланс пищи и энергии для отдельного животного организма можно, таким образом, представить в виде следующего уравнения:
Эп = Эд +Эпр +Эп.в ,
где Эп - энергия потребленной пищи, Эд - энергия дыхания или обеспечения жизнедеятельности организма, включая движение, поддержание температуры тела, сердцебиение и т. п., Эпр - энергия прироста (запасенная в теле организма-потребителя), Эп.в - энергия продуктов выделения (в основном экскрементов).
Количество энергии, расходуемой организмами на различные цели, неоднозначно. В периоды интенсивной жизнедеятельности взрослого организма в теле его может совершенно не фиксироваться энергия. Наоборот, траты ее в ряде случаев превышают поступление (организм теряет вес). В то же время в периоды интенсивного роста организмов, особенно в периоды размножения (беременности), в теле фиксируется значительное количество энергии.
Выделение энергии с экскрементами у плотоядных животных (например, хищников) невелико, у травоядных оно более значительно, а гусеницы некоторых насекомых, питающиеся растениями, выделяют с экскрементами до 70% энергии. Однако при всем разнообразии расходов энергии в среднем максимальны траты на дыхание, которые в сумме с неусвоенной пищей составляют около 90% от потребленной. Поэтому переход энергии с одного трофического уровня на другой в среднем принимается близким к 10% от энергии, потребленной с пищей. Эта закономерность рассматривается обычно как «правило десяти процентов».
Данное правило надо оценивать как относительное, ориентировочное. Вместе с тем из него следует, что цепь питания имеет ограниченное количество уровней, обычно не более 4—5. Пройдя через них, практически вся энергия оказывается рассеянной.
Закономерности потока и рассеивания энергии имеют важные в практическом отношении следствия. Во-первых, с энергетической точки зрения крайне нецелесообразно потребление животной продукции, особенно с высоких уровней цепей питания. Образование этой продукции связано с большими потерями (рассеиванием) энергии. Особенно велики потери энергии при переходе с первого трофического уровня на второй, от растений к травоядным животным.
Часто в экологической литературе рассматривается в качестве примера цепь питания: люцерна-телята-мальчик. Показано, что если бы мальчик весом 48 кг питался только телятиной, то за год ему потребовалось бы для обеспечения жизнедеятельности 4,5 теленка, для питания которых, в свою очередь, необходим урожай люцерны с площади 4 га весом 8211 кг. Такова энергетическая цена животной пищи.
Во-вторых, чтобы сократить вероятность дефицита продуктов питания для интенсивно возрастающей численности населения (по закономерности, близкой к экспоненте), надо, чтобы в рационе людей больший удельный вес занимала растительная пища. Энергетически идеально - вегетарианство.
В-третьих, для увеличения КПД использования пищи при получении животноводческой продукции в условиях культурного хозяйства очень важно уменьшить основную статью нерационального расходования энергии - ее траты на дыхание. Это возможно за счет поддержания оптимального температурного режима в животноводческих помещениях, ограничения подвижности животных и, естественно, сбалансированности кормового рациона по различным элементам питания, а также применения различных биотехнических приемов (умеренные добавки стимуляторов роста, веществ, способствующих улучшению аппетита и т. п.).
Дата добавления: 2016-01-03; просмотров: 1814;