СТАДИИ КЛЕТОЧНОГО ЦИКЛА И ТОЧКИ ЕГО ПРОВЕРКИ
ОПУХОЛИ. ОПРЕДЕЛЕНИЕ ПОНЯТИЯ.
ОТЛИЧИТЕЛЬНЫЕ ОСОБЕННОСТИ ДОБРОКАЧЕСТВЕННЫХ И ЗЛОКАЧЕСТВЕННЫХ ОПУХОЛЕЙ
«Опухоль – это типический патологический процесс, представленный новообразованной тканью, в которой изменения генетического аппарата клеток приводят к нарушению регуляции их роста и дифференцировки(М.А. Пальцев, Н.М. Аничков, 2001). Все опухоли в зависимости от характера роста сгруппированы в два класса: доброкачественные и злокачественные.
| Злокачественные опухоли | Доброкачественные опухоли | |
| 1. | Быстрый рост | Сравнительно медленный рост |
| 2. | Инфильтративный рост | Экспансивный рост |
| 3. | Метастазируют | Не метастазируют |
| 4. | Рецидивируют | Как правило, не рецидивируют |
| 5. | Кахексия | Кахексия не характерна |
| 6. | Иммунодепрессиия | Иммунодепрессия не выражена |
| 7. | Атипизм клеточный и тканевой | Атипизм тканевой |
| 8. | Увеличена площадь капиллярного русла | Сосуды сформированы нормально |
| 9. | Полиморфизм клеток и ядер | Отсутствие полиморфизма |
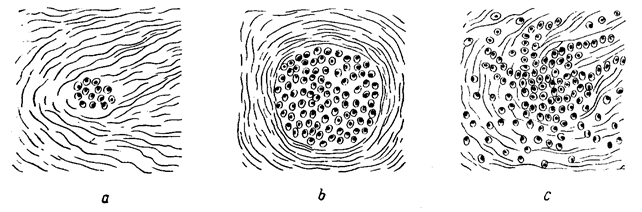
Два типа опухолевого роста (Д.Е. Альперн, 1959)
а - рыхлая соединительная ткань с небольшой группой опухолевых клеток (опухолевый зачаток); b - экспансивный рост; с - инфильтративный рост
Раздел II.
 |
МЕХАНИЗМЫ ГЕНЕТИЧЕСКОГО КОНТРОЛЯ КЛЕТОЧНОГО ЦИКЛА. ИНДУКЦИЯ КЛЕТОЧНОГО ЦИКЛА
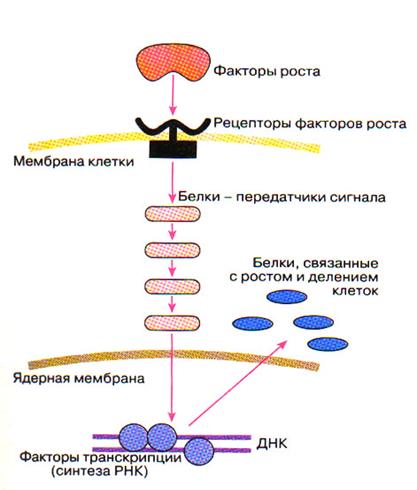
Основные этапы передачи сигнала для роста от мембраны до клеточного ядра
(Г.П. Георгиев, 1999)
· Клетки организма находятся в одном из трех возможных состояний: в цикле, в стадии покоя с сохранением возможности вернуться в цикл и в стадии терминальной дифференцировки, при которой способность делиться полностью утрачена (нейроны головного мозга, сегментоядерные нейтрофилы).
· В норме стимуляция клеточного роста (вхождение клетки в цикл) инициируется факторами роста, которые воспринимаются с помощью соответствующих рецепторов.
· Последующая трансдукция сигнала от активированного рецептора до клеточного ядра осуществляется с участием многих других белков - передатчиков сигнала. Это своего рода клеточное "реле". Такая передача часто идет путем фосфорилирования одним белком второго, вторым - третьего и т.д.
· Цепи передачи сигналов заканчиваются в клеточном ядре. Там происходит активация так называемых факторов транскрипции, т.е. белков, связывающихся с регуляторными участками определенных генов ДНК и активирующих транскрипцию данных генов. Иными словами, под действием факторов транскрипции на соответствующих генах происходит синтез матричных РНК, а на матрице последних - белков. Это те белки, которые нужны для роста и размножения клеток.
· Таким образом, после получения митогенного сигнала в клетке должен произойти ряд событий, включающих в себя сложные преобразования и взамодействия регуляторных молекул, прежде чем она будет в состоянии приступить к репликации ДНК.
· Клеточный цикл в опухолевой ткани насчитывает те же 5 фаз (G0, G1, S, G2 и M), что и в норме. «Вопреки иллюзорному представлению о большей скорости деления малигнизированных клеток современные данные говорят о том, что общая продолжительность их клеточного цикла равна периоду нормального цикла или даже его превышает. Таким образом, рост опухолей не связан с укорочением клеточного цикла» (М.А. Пальцев, Н.М. Аничков, 2001).
СТАДИИ КЛЕТОЧНОГО ЦИКЛА И ТОЧКИ ЕГО ПРОВЕРКИ
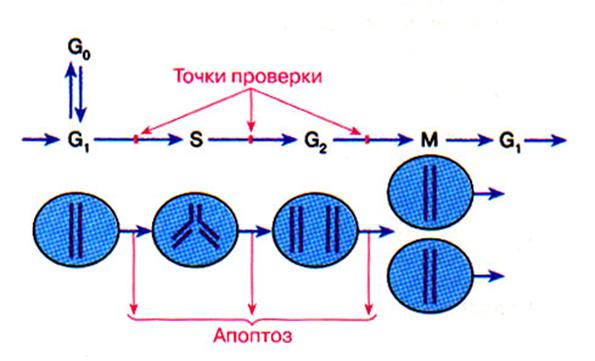 |
Точки проверки клеточного цикла
(Г.П. Георгиев, 1999)
· Наиболее заметными периодами цикла являются митоз (М) и синтез ДНК (фаза S), между которыми выделяются два промежуточных периода (gap) - G1 и G2. На протяжении фазы G1 существует ответственный момент: так называемая точка рестрикции (R), когда решается, войдет ли клетка в следующий цикл деления или предпочтет стадию покоя G0, в которой она может находиться неопределенно долго.
· Вхождение клетки в цикл - процесс вероятностный, определяемый сочетанием ряда внешних и внутренних условий. Однако, после того, как выбор сделан, следующие этапы прохождения по циклу совершаются автоматически. И хотя клетка может быть блокирована на той или иной стадии деления, обычно это является следствием каких-то особых обстоятельств.
· Критически важными в цикле являются моменты вхождения клетки в фазу синтеза ДНК (граница фаз G1/S) и в митоз (граница фаз G2/М), где действуют своеобразные контрольно-пропускные пункты "checkpoints",которые проверяют целостность ДНК (готовность к ее репликации) в первом случае и завершенность репликации - во втором.
 |
РЕГУЛЯЦИЯ КЛЕТОЧНОГО ЦИКЛА. РОЛЬ ЦИКЛИНОВ И ЦИКЛИН-ЗАВИСИМЫХ КИНАЗ
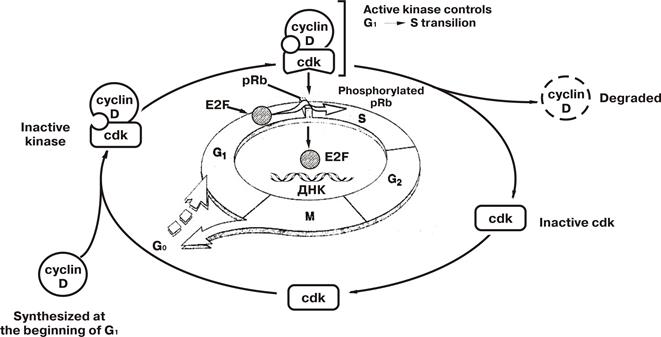
Взаимодействие фактора транскрипции E2F, белка Rb и циклин-зависимых киназ в регуляции клеточного цикла
(по V. Cumar, R.S. Cotran, S. L. Robbins, 1997)
· Для перехода клетки в S-фазу клеточного цикла требуется взаимодействие фактора транскрипции E2F с регуляторными участками генов (гены S-фазы).
· В покоящейся клетке E2F находится в комплексе (связанном состоянии) с протеином Rb (ретинобластомы) и не может осуществить эту функцию. Для этого E2F необходимо высвободить из данного комплекса, для чего требуется фосфорилирование рRb . Такое фосфорилирование обеспечивается циклин-зависимыми киназами (сdk).
· Конститутивно экспрессируясь в неактивном состоянии, cdk в последующем активируются при взаимодействии с циклинами, в частности с циклином D, который синтезируется в начале фазы G1.
· Активные сdk фосфорилируют рRb, который утрачивает связь с E2F, что обеспечивает клетке возможность перехода в S-фазу.
· После вступления клетки в S-фазу циклин D разрушается, а сdk возвращаются в неактивное состояние.
Дата добавления: 2015-12-29; просмотров: 1364;