Биотические факторы
Все организмы, независимо от их видовой принадлежности, не только испытывают влияние среды, но и сами прямым образом или косвенно влияют на нее. Поэтому среди биотических факторов различают фитогенные, зоогенные и антропогенные факторы.
Фитогенное воздействие на среду чрезвычайно многообразно. Назовем лишь некоторые примеры и, прежде всего, фотосинтез, который определяет газовый состав среды. В результате фотосинтеза зеленые растения выделяют в атмосферу кислород. Они же и поглощают углекислый газ. Наземные растения оказывают влияние на среду также путем изменения структуры и состава почвы. Далее, азотфиксирующие бактерии, извлекая азот атмосферы, также влияют на ее газовый состав.
Зоогенные воздействия на среду также многообразны. Например, потребляя для дыхания кислород, животные также изменяют газовый состав атмосферы. Животные, обитающие в почве, оказывают значительное влияние на ее механический состав.
Антропогенные воздействия мы рассмотрим в гл. XVIII.
Все организмы находятся между собой в самых различных отношениях, которые, в частности, проявляются в форме симбиоза (мутуализма, комменсализма, хищничества, паразитизма), конкуренции и антагонизма.
Симбиоз (от греч. symbiosis — сожительство) — это очень распространенная форма во взаимоотношениях организмов, принадлежащих к разным видам. В рамках симбиоза различают мутуализм, комменсализм, хищничество и паразитизм.
Мутуализм — это взаимодействие между двумя организмами разных видов, которое выгодно для каждого из них. Например, азотфиксирующие клубеньковые бактерии обитают на корнях бобовых растений, конвертируя атмосферный азот в форму, доступную для усвоения этими растениями. Следовательно, бактерии обеспечивают растения азотом. В свою очередь растения обеспечивают клубеньковые бактерии всеми необходимыми питательными веществами. Мутуализмом можно считать также взаимодействие между микроорганизмами, обитающими в толстом отделе кишечника человека, и самим человеком. Для микроорганизмов выгода определяется тем, что они обеспечивают свои питательные потребности за счет содержимого кишечника, а для человека выгода состоит в том, что микроорганизмы осуществляют дополнительное переваривание пищи и еще синтезируют крайне необходимый для него витамин К. В мире цветковых растений мутуализмом является опыление насекомыми растений и питание насекомых нектаром растений. Мутуализм значим и в «переработке» органических веществ. Например, переваривание целлюлозы в желудке (рубце) крупного рогатого скота обеспечивается содержащимися в нем бактериями.
Комменсализм — это межвидовое взаимодействие между организмами, при котором один организм получает выгоду за счет другого, не повреждая его, тогда как другой организм от этого взаимодействия не имеет ни выгоды, ни ущерба. Например, некоторые виды морских полипов поселяются на поверхности тела крупных рыб, питаясь их выделениями, но для рыб данное сожительство является индифферентным, т. е. не имеет никакого значения.
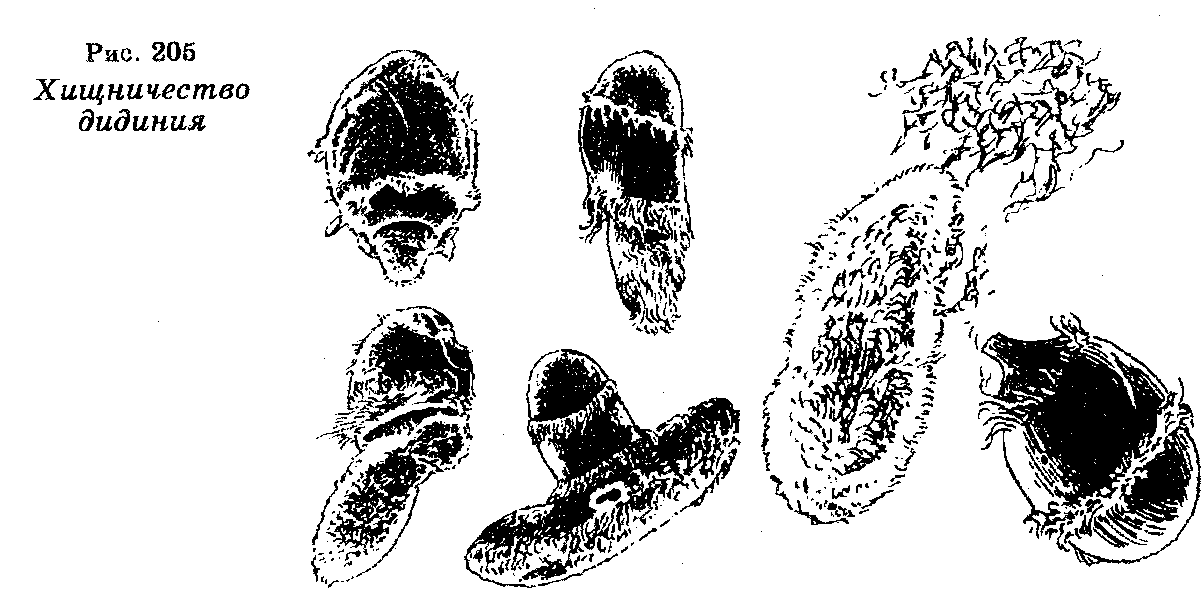
Хищничество — это особый способ жизни, при котором один организм (хищник) живет за счет другого (жертвы), убивая его. Хищничество наблюдается уже у простейших. Например, известна инфузория дидиний (Didinium nasutum), которая является хищником для других простейших, в частности, парамеций. Плавая в воде, дидиний парализует парамецию, прикрепляется к ней, а затем заглатывает. Процесс переваривания одиночной парамеции составляет 2 часа (рис. 205).
Примеры хищничества высших животных в природе многочисленны. Убивая и поедая жертвы, хищники приносят вред популяциям организмов-жерт.в. Но хищничество иногда оказывается и полезным. Например, волки, поедая ослабленных (больных) лосей, способствуют оздоровлению лосиных популяций.
Уменьшение численности хищников может привести к резкому возрастанию численности жертв. Иногда имеет место синхронный рост численности и хищников и жертв. Например, в северных широтах эта синхронность отмечается каждые несколько лет между численностями леммингов, мышей, с одной стороны, и питающихся ими песцов, лис и полярных сов, с другой.
Паразитизм — это форма взаимоотношений организмов, при • которой один (паразит) живет за счет другого (хозяина). Как правило, хозяину присущи большие размеры тела, чем паразиту. В отличие от хищников паразиты не должны убивать хозяев, иначе тем самым они убьют и себя. Поэтому паразиты вызывают болезнь хозяина, но смерть хозяина от этой болезни бывает лишь в очень редких случаях.
Паразитизм очень широко распространен в природе, но его следует отличать от сапробиоза, при котором организм питается мертвым материалом другого организма, хотя некоторые сапро-бионты иногда могут стать паразитами. С эволюционной точки зрения наиболее результативными являются те паразиты, которые вызывают незначительные повреждения хозяина
В процессе эволюции паразитизма паразиты приобрели исключительную специализацию. Они потеряли, прежде всего, те органы, которые в условиях паразитизма перестали быть ценными для их существования. Напротив, у них развились новые органы, которые обеспечивают их способность существовать за счет хозяев, прикрепляясь к его клеткам, обитая в его полостях и жидкостях.
Среди морфологических и биологически специализированных модификаций паразитов наибольшую важность представляет способность к физиологической адаптации в организме хозяина и наличие сложных жизненных циклов, которые обеспечивают распространение их среди новых хозяев. Физиологическая адаптация к хозяину характеризуется хозяйской специфичностью, т. е. паразит может существовать лишь в хозяевах, принадлежащих к определенным видам. Иногда диапазон хозяйской специфичности является очень широким, в результате чего паразит может паразитировать на организмах многих видов, причем родственные виды хозяев иногда имеют и родственных паразитов. Однако часто хозяйская специфичность настолько узка, что паразит может использовать в качестве хозяина лишь организмы одного единственного вида.
Предполагают, что эволюция видов паразитов и их хозяев шла синхронно (правило Фаренгольца). Хозяева оказывают влияние на эволюции паразитов, но и паразиты влияют на эволюцию хозяев. Например, резистентность к малярии, вызываемой в Западной Африке Р. falciparum, связана с тем, что этот паразит является причиной появления новых аллелей в комплексе гистонесовместимос-ти аборигенов.
Хозяев паразитов классифицируют на дефинитивных и промежуточных.
Организм-хозяин, в котором паразит приобретает половую зрелость, является дефинитивным (постоянным) хозяином, а организм-хозяин, в котором паразит проходит лишь некоторые стадии своего развития, служит промежуточным хозяином. Чтобы облегчить собственный перенос от одного хозяина к другому, паразиты часто используют промежуточных хозяев. Иногда завершение жизненного цикла паразита связано с последовательной сменой нескольких промежуточных хозяев.
Распространение паразитов определяется, прежде всего, географическим распространением их хозяев и переносчиков, а также климатическими условиями, в которых обитают их хозяева. Поскольку многие паразиты могут переходить от хозяина к хозяину, то онтогенез паразитов связан с модификацией их репродуктивной системы в направлении повышения ее мощности. В результате этого у паразитов развилась способность продуцировать потомство в огромных количествах. Повышение количества потомства часто связано с бесполым размножением, гермафродитизмом.
Паразитов классифицируют на временных (комары, мухи-жигалки, блохи и др.) и постоянных (простейшие, гельминты и др.), на эндопаразитов (простейшие, гельминты) и эктопаразитов (вши, блохи и др.). Паразитизм имеет важное значение в сельском хозяйстве и медицине, так как многие паразиты являются возбудителями болезней животных и человека.
Конкуренция — это взаимоотношение между организмами при использовании одинаковых жизненных ресурсов (пищи, света, территории и т. д.), которых, однако, недостаточно в определенных средах при значительном росте популяций. Поэтому она ведет к снижению плодовитости, повышению смертности и к другим неблагоприятным последствиям. Конкуренция бывает как внутривидовой, так и межвидовой и сопровождается она борьбой за существование. Внутривидовая конкуренция является более ожесточенной, поскольку организмы нуждаются в одних и тех же ресурсах. Голод иногда у отдельных видов ведет к каннибализму. Конкуренция и борьба за существование играют большую роль в ограничении численности популяции. Их следствием является естественный отбор (см. гл. XIV).
Антагонизм — это форма взаимоотношений, когда один вид в процессе жизни угнетает другой. Например, известны виды растений, которые выделяют биологически активные вещества (фитон-циды), угнетающие развитие растений других видов. Формой антагонизма является антибиоз, который имеет место в случае бактерий и микроскопических грибов, когда организмы одного вида подавляют рост организмов другого вида или сразу нескольких видов. Эта подавляющая способность зависит от продукции организмами антибиотических видов веществ, получивших название антибиотиков. Наиболее известными антибиотиками являются ампициллин, стрептомицин, тетрациклин, хлорамфеникол и другие.
§ 96 Факторы защиты организма (иммунитет)
В ходе эволюции в процессе взаимодействия между собой животные организмы выработали многие факторы собственной защиты от проникновения в них вирусов, бактерий, простейших и других патогенных факторов, которые действуют либо механически, либо вырабатывают токсины, оказывающие неблагоприятное влияние на клетки и ткани хозяина.
Факторами естественной защиты человека являются кожа, поверхностные структуры (слизистые оболочки) дыхательного тракта, соляная кислота в желудке, лизопим, интерферон, белые форменные элементы крови и антитела. Защитная функция кожи, слизистых оболочек дыхательного тракта, соляной кислоты, лизоцима, пропердина и интерферона заключается в том, что они препятствуют проникновению патогенов в организм, действуя бактерицидно. Однако если патогены все же проникают в организм, тогда вступает в действие и иммунная система посредством механизмов иммунитета в виде фагоцитарной и гуморальной защиты.
Иммунитет (от лат. immunitas — освобождение от чего-либо) — это невосприимчивость организма к болезнетворным агентам, продуктам их жизнедеятельности и к генетически чужеродным веществам, обладающим антигенными свойствами. Можно сказать, что иммунитет представляет собой способность организма отличать чужеродный материал от своего, например, чужеродный белок от своего и нейтрализовать этот материал. В отличие от наследственности, которая охраняет организмы от резких изменений на протяжении поколений, иммунитет осуществляет охрану на протяжении индивидуальной жизни организма (онтогенеза).
Охранительная роль иммунитета распространяется не только на вирусы, простейшие, грибы, гельминты, но и на чужеродные трансплантанты тканей и органов. Она распространяется также на аутоиммунные процессы, возникающие в организме. Например, в механизме возникновения сахарного диабета у человека имеют значение аутоиммунные процессы против белков, содержащихся в клетках островков Лангерганса панкреатической железы.
Различают иммунитет инфекционный и неинфекционный.
Инфекционный иммунитет классифицируют на противовирусный, антимикробный (антибактериальный) и антитоксический. В случае противовирусного иммунитета (при гриппе, полиомиелите человека и других вирусных инфекциях) происходит разрушение вирусных частиц, при антимикробном иммунитете (при дизентерии человека, бруцеллезе человека и животных) происходит обезвреживание бактериального возбудителя, тогда как при антитоксическом (при столбняке, ботулизме человека и животных и др.) имеет место разрушение токсина, продуцируемого микробами в организме. В рамках инфекционного иммунитета различают также врожденный и приобретенный иммунитет. Врожденный иммунитет — это естественный иммунитет, возникший в ходе исторического развития организмов и передающийся по наследству. Например, куры не восприимчивы к возбудителю сибирской язвы, причем эта невосприимчивость передается по наследству от одного поколения к другому. Приобретенный иммунитет приобретается организмом в ходе его жизни. Такой иммунитет классифицируют на активный и пассивный иммунитет. Активный иммунитет возникает после перенесения организмами болезни или после их вакцинации, тогда как пассивный — после введения в организм готовых антител (сыворотки, содержащей антитела).
Неинфекционный иммунитет является результатом исторически сложившейся генетической несовместимости. Например, неинфекционным иммунитетом является несовместимость донора и реципиента по группам крови, проявляющаяся в виде тяжелых осложнений при переливании несовместимой крови. Неинфекционным иммунитетом является также трансплантационный иммунитет, развивающийся при пересадке сердца и других органов у человека. Трансплантационный иммунитет возникает, когда ткани донора и реципиента неидентичны. Этот иммунитет проявляется в виде разрушения (отторжения) пересаженной ткани или органа уже через 8-23 дня после пересадки.
Различают клеточный, гуморальный и тканевой иммунитет.
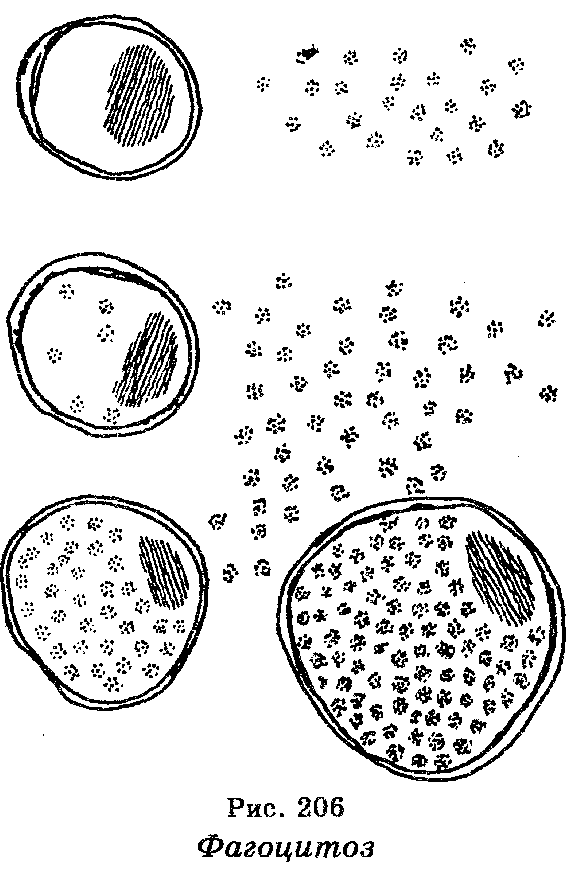
Клеточный иммунитет заключается в фагоцитозе, т. е. в захва-тывании и переваривании фагоцитами (специализированными клетками) бактерий, проникших в организм (рис. 206). Фагоцитами являются клетки белой крови, называемые микрофагами (эозинофилы, нейтрофилы и базофилы) и макрофаги (подвижные клетки крови — моноциты, клетки лимфатических узлов и селезенки, эндотелий кровеносных сосудов).
 Гуморальный иммунитет — это иммунитет, связанный с выработкой антител и обусловленный взаимодействием антигенов и антител. Антигены — это чужеродные для организма вещества. Ими являются белки, липопротеи-ды, белки в соединении с полисаха-ридами, нуклеиновые кислоты. Антигенами являются также ферменты, токсины, яды змей и пауков, вирусы, бактерии, простейшие. Характерными свойствами антигенов являются их чужеродность и специфичность (видовая, групповая, органная и тканевая), а также то, что они вызывают иммунный ответ в виде образования антител. Можно сказать, что антигены выполняют роль генераторов антител. Известны также аутоантигены, образующиеся в самом организме в результате повреждения тканей. Например, аутоантигены, связанные с множественным склерозом человека, присутствуют в миелиновой оболочке, окружающей нервные волокна центральной нервной системы, представляя собой белок р-кристаллин.
Гуморальный иммунитет — это иммунитет, связанный с выработкой антител и обусловленный взаимодействием антигенов и антител. Антигены — это чужеродные для организма вещества. Ими являются белки, липопротеи-ды, белки в соединении с полисаха-ридами, нуклеиновые кислоты. Антигенами являются также ферменты, токсины, яды змей и пауков, вирусы, бактерии, простейшие. Характерными свойствами антигенов являются их чужеродность и специфичность (видовая, групповая, органная и тканевая), а также то, что они вызывают иммунный ответ в виде образования антител. Можно сказать, что антигены выполняют роль генераторов антител. Известны также аутоантигены, образующиеся в самом организме в результате повреждения тканей. Например, аутоантигены, связанные с множественным склерозом человека, присутствуют в миелиновой оболочке, окружающей нервные волокна центральной нервной системы, представляя собой белок р-кристаллин.
Антитела — это белки, в частности, иммуноглобулин, содержащийся в сыворотке крови. Известно около десяти групп разных антител, среди которых у человека наиболее часто встречаемыми являются антитела групп IgG, IgM, I^D, IgE и IgA. Иммуноглобулины (антитела) каждой из этих групп состоят из полипептидных цепей разной молекулярной массы. Например, антитела IgG состоят из четырех полипептидных цепей, две из которых имеют молекулярную массу 22 500, а две другие — 50 000. В соответствии со способом действия антител на антигены различают нейтрализующие, лизирующие, коагулирующие и способствующие фагоциту антитела. Помимо антител, циркулирующих в плазме крови, существуют клеточные антитела, связанные с поверхностью клетки.
В ходе эволюции выработался комплекс между способностью животных отвечать немедленно на какой-либо один из миллионов антигенов и «мощностью» иммунитета. Молодые млекопитающие животные способны распознавать и отвечать на внедрение в организм многих антигенов, но эта их способность является небольшой. Способность животных отвечать на антиген быстрее и мощнее возникает лишь со временем. Однако иммунная система может быть мобилизована до того, как чужеродный фактор попал в организм. Эту особенность используют в иммунизации человека и животных вакцинами, содержащими антигены возбудителей тех или иных инфекций с целью профилактики против этих инфекций. Иногда для увеличения продукции специфических антител используют двухтрехкратную вакцинацию в интервалы через несколько недель.
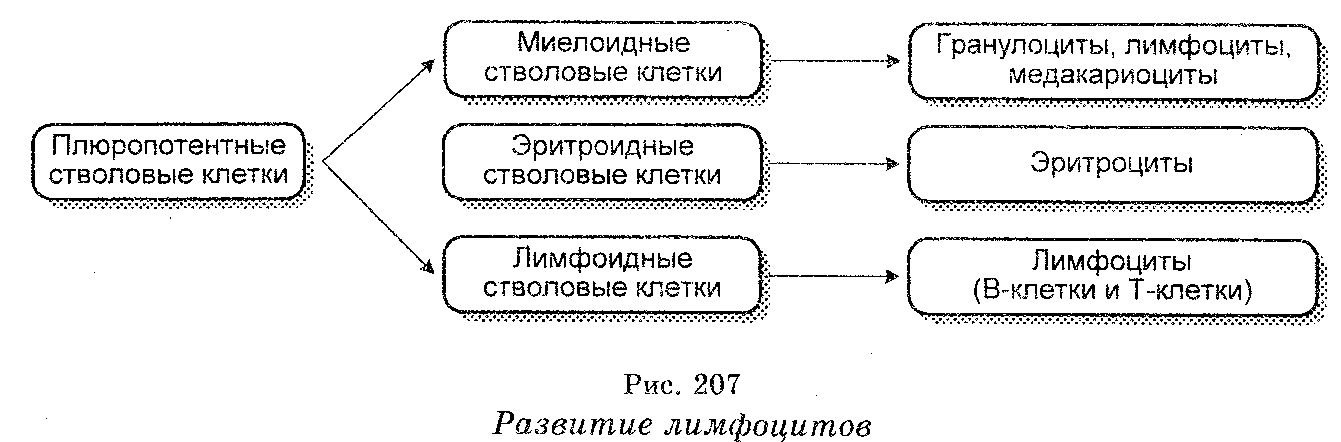
Появление антител отражает одну сторону иммунного ответа, тогда как другая сторона связана с включением в «работу» лимфоцитов, поскольку морфологическим субстратом антител являются лимфоидные клетки лимфоузлов, селезенки, костного мозга, миндалин, тимуса, а за развитие иммунного процесса и за его специфичность ответственны лимфоциты, плазматические клетки и моноциты (макрофаги) и поскольку центральная роль в иммунном ответе принадлежит лимфоцитам. Они развиваются из так называемых плюропотентных стволовых клеток, которые являются родоначальниками всех клеток крови (эритроцитов, лейкоцитов и тромбоцитов).
Различают долгоживущие тимусзависимые Т-лимфоциты и ко-роткоживущие В-лимфоциты (рис. 207). Т-лимфоциты развиваются в тимусе, тогда как В-лимфоциты — в костном мозге взрослых или в печени плодов. В тимусе Т-клетки развиваются из клеток-предшественников, поступающих в тимус из кроветворных органов. В-клетки у млекопитающих возникают непосредственно из стволовых клеток в кроветворных органах, а у птиц — в фабрициевой бурсе (сумке) из клеток-предшественников, поступающих в бурсу с кровью из кроветворных тканей. Кроветворные ткани, тимус и фабрициеву бурсу называют первичными лимфоидными органами.
После дифференцировки в первичном лимфоидном органе часть лимфоцитов с током крови переносится во вторичные лимфоидные органы (лимфатические узлы, селезенка, аппендикс, миндалины, аденоиды и пейеровы бляшки тонкого кишечника). Именно здесь Т-клетки и В-клетки реагируют с антигенами. Т-лимфоциты первоначально распознают чужеродный антиген, а затем становятся хранителями иммунологической памяти и переносчиками этой информации антителообразующими клетками. В-лимфоциты образуются в огромном количестве (ежедневно по нескольку миллионов). Они активируются Т-клетками и дифференцируются или трансформируются в плазматические клетки, непосредственно образующие антитела (растворимые Иммуноглобулины) против распознанных антигенов.
Лимфоциты — это иммунокомпетентные клетки. Они способны переносить в другой организм свойства иммунного организма — противотканевый иммунитет, иммунологическую память, аллергическую активность. Лимфоциты Т и В осуществляют иммунологи-ческий надзор и память в отношении чужеродного антигена. Реализация гуморального иммунного ответа связана с В-лимфоцитами, которые становятся продуцентами антител, а клеточного — с Т-лим-фоцитами, причем в обоих случаях с участием макрофагов.
Почему аутоантитела не распознают собственные макромолекулы? Считают, что распознание аутоантителами собственных макромолекул не происходит по причине гибели лимфох^итов, выполняющих эту функцию. Несомненно, что данное явление имеет место после развития иммунной системы организма, но до встречи организма с чужеродными макромолекулами.
Иммунный ответ организма координирует белки — цитокины, которые являются своеобразными иммунологическими гормонами.
Антитела, возникшие в результате иммунизации, называют иммунными, тогда как в сыворотках крови нормальных людей и животных могут находиться их естественные нормальные антитела. Схема развития лимфоцитов является относительно простой. Предполагают, что в ходе дробления оплодотворенной яйцеклетки возникает одна клетка, которая является предшественником клеток Т и В. Последующие деления этой клетки продуцирует только клетки Т и В, потомство каждой из которых развивает способность распознавать только один антиген (одни макромолекулы). Однако некоторые из этих антигенов являются собственными макромолекулами организмов. Лимфоциты, распознающие собственные антигены, погибают или делаются «нераспознающими», причем время, когда это происходит, зависит от вида организмов. Например, у мышей это происходит в период рождения, а у человека — еще в период эмбрионального развития.
Известно также, что при размножении в клетках нервной системы вирусов полиомиелита, клещевого энцефалита и др. образуются антигены, вызывающие синтез аутоантител. Эти реакции аутоантителообразования ведут к аутоиммунным болезням (гломерулонефриту, некоторым гемолитическим анемиям, множественному склерозу человека).
Известно, что организм животных обладает способностью синтезировать гигантское разнообразие антител. Например, мыши могут синтезировать антитела около 2´106 специфических типов. Исходя из того, что каждый антиген кодируется только одним геном, а размер одного гена составляет примерно 1000 нуклеотидов, часть мышиного генома, ответственного за синтез антигенов, равна 2´109 нуклеотидным парам.
Для объяснения механизмов образования антител предложено несколько теорий, из которых наиболее популярной является «кло-нально-селекционная» теория (Ф. Вернет, 1961). В соответствии с этой теорией в организме имеется исключительно многочисленная популяция лимфоидных клеток, порядка 1 х 10". Эта популяция очень гетерогенна генетически, в результате чего каждая группа лимфоцитов обладает разным родством к антигену. Вследствие контакта с антигеном лимфоциты, обладающие на своей поверхности белками-рецепторами антигена, подвергаются пролиферации и трансформации в плазматические клетки, продуцирующие антитела. Таким образом, антиген избирательно стимулирует те клетки, у которых к нему есть рецепторы. Давая затем потомство, они образуют множество клонов, состоящих из Т-клеток или В-клеток и обладающих одинаковой антигенной специфичностью.
Клетки определенных клонов лимфоцитов могут сохранять память об антигенном стимуле в течение нескольких делений. Это носит название иммунологической памяти., и на этом явлении основано длительное поддержание иммунитета.
Поскольку апоптоз клеток сопровождается экскрецией в межклеточные пространства деградированной ДЦК итистонов, то считают, что последние являются аутоантигенами, определяя аутоим-мунные реакции.
Иногда иммунная система не способна реагировать на собственные антигены. Это называют естественной иммунологической толерантностью. Если же толерантность к собственным антигенам нарушится, то результатом будет развитие аутоиммунологических болезней. Избыточная антигенная стимуляция приводит к открытому в 1953 г. австралийским ученым П. Медаваром и чешским ученым М. Гашеком явлению приобретенной иммунологической толерантности (от лат. tolerantia — терпение).
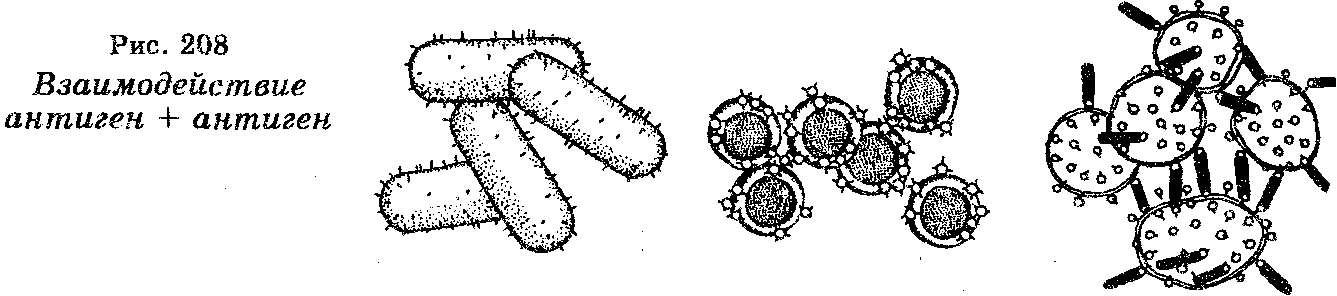
Как уже отмечено, антитела представляют собой белки-имму-ноглобулины. Количество их в плазме крови составляет примерно 20% всех плазменных белков. Для них характерно наличие анти-генсвязывающих участков, причем каждый клон В-клеток продуцирует антитела со специфическими антигенсвязывающими участками. Эти участки принимают участие во взаимодействии антигена с антителом (рис. 208).
Основными реакциями иммунитета являются нейтрализация токсинов (антитоксинами), преципитация бактерий (преципитинами), агглютинация бактерий (агглютининами), лизис бактерий (лизина-ми), связывание комплемента, опсонизация (опсонинами). Эти естественные реакции иммунитета широко используют в лабораториях для диагностики многих болезней человека и домашних животных.
Иммунный ответ клеточного типа детерминируется реакциями Т-клеток, которые у позвоночных чрезвычайно важны в защите от вирусов и грибов. Различают цитотоксические Т-клетки, убивающие соматические клетки, инфицированные вирусом, Т-хелперы, помогающие В-лимфоцитам в образовании антител, и Т-супрессоры, подавляющие иммунные реакции. Считают, что Т-хелперы и Т-супрессоры являются главными регуляторами иммунных ответов организмов.
Тканевой иммунитет у животных и человека обеспечивается кожей, слизистыми оболочками, лимфатическими узлами, тканями мышц, мускулатуры кишечника и матки, сывороткой крови и другими жидкостями. Этот иммунитет является неспецифическим.
Часто иммунологическая реактивность организмов может изменяться. У человека и животных различают явление, называемое аллергией. Аллергия (от греч. allos — другой, ergon — действие) — это измененная реактивность организма, наступающая под влиянием микробов, токсинов, лечебных препаратов и других веществ, называемых аллергенами. Аллергенами могут служить пыль подушек, перхоть собак, кошек, лошадей, пыль шерсти, хлопка, пыльца растений, яйца, кофе, цитрусовые и др. Как правило, она возникает в результате повторного введения или попадания в организм того или иного аллергена.
Аллергия проявляется либо очень быстро в форме анафилаксии (судороги, выделение мочи и кала, повышение температуры), сывороточной болезни и острого ревматизма либо в замедленной форме (лекарственная болезнь, вызываемая антибиотиками и другими препаратами).
Аллергические, инфекционные и неинфекционные болезни с выраженным проявлением воспалительных реакций, а также иммуно-дефицитные состояния и аутоиммунные заболевания человека в настоящее время довольно широко распространены. Считают, что в основе иммунологических нарушений лежит ряд причин, а именно:
1. Снижение эффективности естественного отбора на устойчивость к инфекциям, т. е. снижение роли отбора в совершенствовании иммунной системы организмов, включая человека.
2. Снижение стимуляции лимфоцитов Т и В, равно как и клеток иммунной системы, антигенами бактерий, обитающих в кишечнике и других полостях тела животных и человека, из-за широкого использования антибиотиков, которые убивают бактерий.
3. Интенсификация воздействия на животных и человека экологических факторов, обладающих иммунологическим эффектом. Иммунитет является недостаточным либо совершенно отсутствует у животных и человека против гельминтов.
§ 97 Пространство, местообитания,
Биомы, сообщства
Жизнь сосредоточена в океанах (морях) и на материках. Однако пространство океанов и материков измеряется очень большими расстояниями, на которых могут существовать совершенно различные живые формы. Поэтому в целях упорядочивания пространство океанов и материков районируют, выделяя фаунистические и флористические области.
В океане различают 10 фаунистических (зоогеографических) областей, в частности. Арктическую, Атлантическую, Тихоокеаническую, Западноатлантическую, Восточноатлантическую, Магелланову и другие. Что касается материков, то на них различают таких 6-9 областей. По одной из классификаций выделяют Палеаркти-ческую (Европа, Средняя Азия, Северная Азия и Северная Африка), Неоарктическую (Северная Америка), Кругоантарктическую (Антарктика), Неотропическую (южная Америка), Эфиопскую (Африка), Восточную (Юго-Восточная Азия) и Австралийскую области. Для каждой из этих областей характерна. своя фауна. Существуют и другие классификации материковых фаунистических областей, но они не имеют особых отличий от приведенной классификации.
Выделение флористических областей на пространствах морей и материков имеет некоторое отличие, т. к. ботаники предпочитают применять термин «флора» (на практике) лишь к сосудистым растениям. В случае низших растений чаще используют такие словосочетания, как биофлора, альгофлора и т. д. Поэтому флористическое районирование распространяется практически лишь на материки, на территориях которых выделяет ряд областей (фитохорионов) с присущими им определенным видовым составом растений. В выделении областей учитывают эндемизм растений, т. е. приуроченность видов, родов и других систематических единиц растений к определенным территориям, границы которых иногда бывают несколько размытыми. Растения видов, родов, семейств и других систематических единиц, приуроченные в своем распространении к определенным территориям, являются эндемиками.
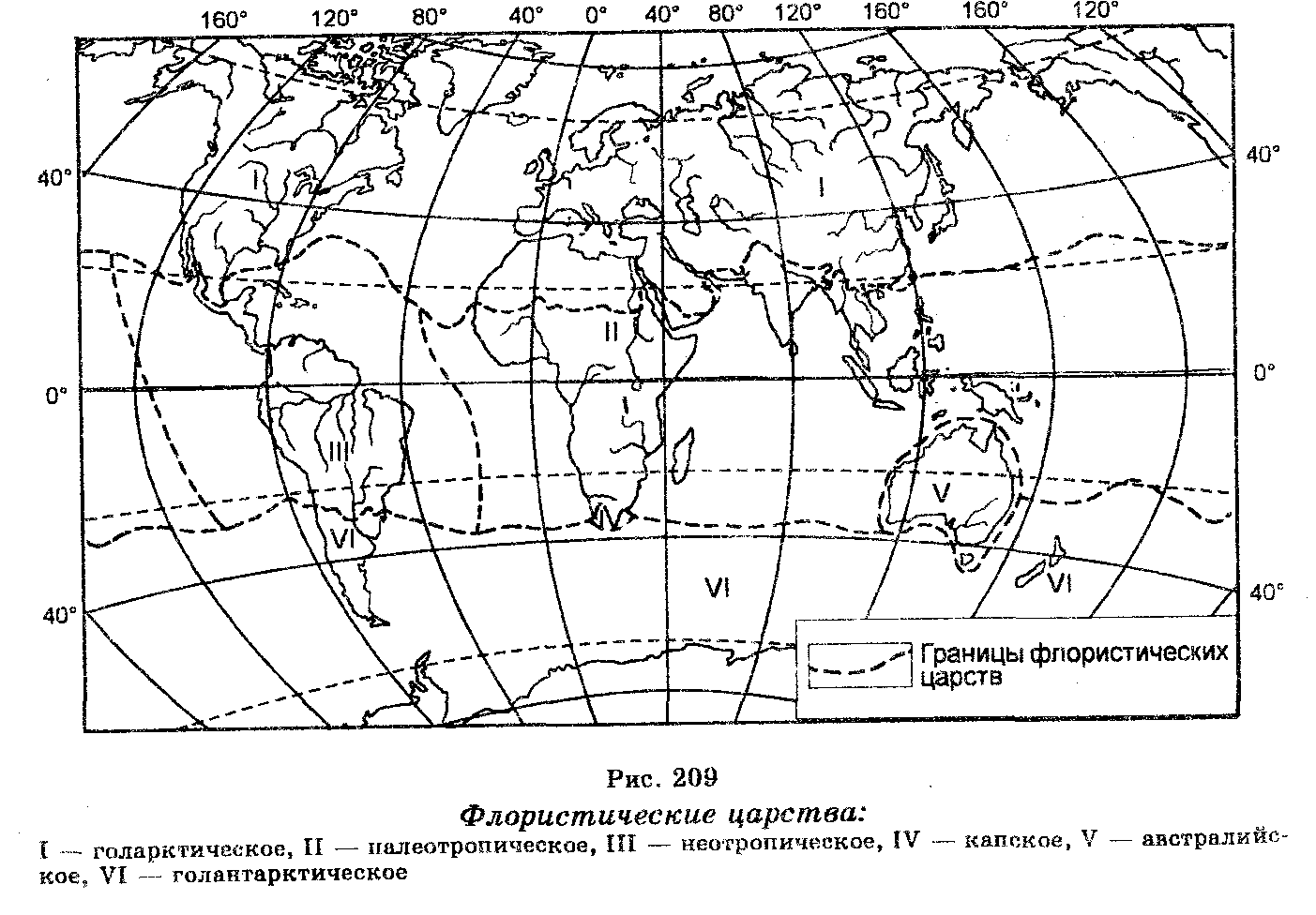
В районировании различают прежде всего флористические царства, которые подразделяют на флористические области, флористические области — на флористические провинции, а последние — на флористические округа.
В соответствии с наиболее распространенной флористической классификацией различают голарктическое, палеотропическое, неотропическое, капское, австралийское и голантарктическое царства (рис. 209). Наша страна укладывается в границы Голарктического царства, которое охватывает Европу, Северную Африку, Северную Америку и ту часть Азии, которая не является тропической. Голарктическая флора представлена такими семействами растений, как лютиковые, гвоздичные, барбарисовые, березовые, ивовые, магнолиевые, розоцветные, крестоцветные и др.
Видовой состав разных флор очень различен. Наиболее богатыми по видовому составу являются флоры тропических широт. Если в Южной Америке насчитывают около 40 тыс видов растений, то в Гренландии их лишь около 400, а на Шпицбергене около 130. В тропических широтах преобладают орхидные (в основном во влажных тропических лесах), молочайные, мареновые, злаки, бобовые, осоковые. В зонах умеренного климата преобладают сложноцветные, бобовые, розоцветные, злаки, осоковые и крестоцветные. Для засушливых районов характерны обычно мареновые.
Местообитание — это, по существу, место, где живет тот или иной организм. Им может быть участок поверхности земли, водоема или воздушного пространства. Размеры местообитания чрезвычайно варьируют, что зависит от вида организма. Например, ме-стообитанием термитов может служить нижняя поверхность гниющего бревна, тогда как местообитанием лососевых является почти весь Тихий океан.
Различают наземные, воздушные и водные местообитания. Наземные местообитания представляют собой поверхность и поверхностные слои Земли. Наиболее богатой является наземная флора тропических широт. Воздушные местообитания являются временными, т. к. все летающие животные являются все же наземными организмами- Наиболее объемными и населенными являются водные местообитания, которые делят на морские и пресноводные.
Морские местообитания занимают около 78% поверхности земного шара и представляют собой слои воды глубиной до 4000 м, в которых еще активна жизнь. Моря и океаны неоднородны по физическим свойствам, и это ведет к тому, что в разных районах Мирового океана существуют и разные температуры. Температуpa морей зависит от места моря, глубины, течения и времени года. Если в полярных и тропических морях она более или менее постоянна, то в морях континентальной зоны она непостоянна, особенно в разные времена года. На глубинах более 2000 м она всегда и везде составляет около 2°С. Между тем, температура имеет ведущее значение в определении зональной распространенности организмов.
Соленость морской воды также ограничивает распространение живых форм. Например, в Мертвом море из-за высокой концентрации солей вообще нет живых организмов.
Свет чрезвычайно важен для организмов, но на глубинах 2000 м и более устанавливается сплошная темнота. Первичные производители органического вещества в море сосредоточены в прибрежных областях и представлены фитопланктоном, который состоит в основном из диатомовых водорослей и динофлагеллят. Но в открытых тропических морях планктона очень мало. Зоопланктон встречается во всех областях моря и представлен мелкими членистоногими, а также яйцами и личинками разных животных.
Установлено, что основные морские сообщества организмов формируются в зависимости от температуры, глубины и солености того или иного моря. Их классифицируют на тропические и полярные морские сообщества, хотя границы между этими сообществами чрезвычайно размыты.
Пресноводные местообитания составляют 2% от всей площади суши. Они представлены реками, озерами, прудами, водохранилищами. Местообитания организмов здесь ограничиваются температурой, мутностью воды, течениями, кислородом и углекислым ва-зом, солями и осмотическим давлением. Наиболее богатыми по видовому содержанию животных и растений являются прибрежные зоны.
Фитопланктон пресных водоемов представлен зелеными и диа-томовыми водорослями, а также цианобактериями.
Растения, животные, физический субстрат и климат находятся в постоянном взаимодействии между собой. В результате этого в местах обитания формируются крупные комплексы (совокупности) организмов, получившие название биомов.
Различают наземные, морские и пресноводные биомы. Каждый из наземных биомов характеризуется определенным набором растительности (деревья, кустарники, травы и т. д.), с которой связаны живущие в этих местообитаниях животные. Биомами являются тундра, хвойные леса, лиственные леса, влажный тропический лес, степь, чаппараль, пустыня.
В одном биоме может обитать одно или несколько сообществ животных и растений, причем видовое разнообразие сообществ зависит от его возраста, климата, продуктивности и др.
Для биомов, населяющих местообитания, характерна определенная климаксная растительность и определенные виды животных. Между обоими компонентами (группировками) биома существует тесная связь.
Эти группировки растений и животных получили название сообществ. Часто сообщества называют по большинству встречающихся в нем организмов.
Размеры сообществ очень колеблются, тем не менее их структура характеризуется упорядоченностью, которая выражается в определенных отношениях между видами, одни из которых встречаются чаще, другие реже. С другой стороны, для сообществ присуща пространственная структура (размещение в пространстве) и видовая. На состав сообществ влияют пищевые связи. Например, использование степи в качестве пастбища сопровождается ростом той же растительности. Но прекращение выпасов заканчивается ростом только ковыля.
Сообщества состоят из популяций организмов разной видовой принадлежности. Тропические сообщества характеризуются большим количеством видов, но численность видов является не очень большой.
Напротив, сообщества полярных морей характеризуются меньшим количеством видов, но численность их является большей.
Например, в море Лаптевых встречается 400 видов животных, в Карском — 1200 видов, в Баренцевом — 2500 видов, в Северном — 3000-4000 видов, в Средиземном — 6000-7000 видов, в Черном — 1200 видов, в Белом — 1000 видов.
Видовой состав сообществ может быть разным, но некоторые виды в них имеют сходные фенотипы и функционально одинаковы. Эти виды называют экологическими двойниками (экологическим эквивалентами).
Облик современных сообществ является результатом эволюции. В свою очередь сообщества влияют на эволюцию видов.
Популяции
Сообщества организмов состоят из видов, а виды — из популяций (см. гл. XIV). Вид может состоять из одной или более популяций, причем разной численности. Между популяциями практически не бывает резких разграничении, хотя степень их изоляции зависит от географических условий, от местообитания, от численности соседних популяций. В результате интенсивного размножения границы между популяциями вообще могут размываться. В случае многих млекопитающих популяции имеют стадный характер.
Что же касается структуры самих популяций, то она может быть пространственной, возрастной, половой и генетической.
Пространственная структура определяется размещением особей в пространстве и зависит от биологических свойств вида, от характера местообитания, от времени года. Например, организмы многих видов предпочитают держаться стаями (птицы) либо стадами (млекопитающие).
Возрастная структура популяции определяется количеством в популяции организмов того или иного возраста, а половая — количеством самцов и самок.
Генетическая структура является отражением количественных отношений между разными генотипами в популяции.
Популяции в сообществах взаимодействуют между собой. Одни из них создают среду для других организмов. Например, деревья служат местом для гнездовий птиц или, затеняя почву, мешают развитию травянистой растительности. Это так называемые типические связи. Но часто организмы связаны между собой пищевыми связями, когда один организм служит пищей для других. Например, лисы используют в пищу мышей-полевок, что и определяет их довольно тесную связь.
Для популяций характерна такая величина, как численность или поголовье. Эта величина не является постоянной, ибо зависит от очень многих факторов (темпов размножения, гибели особей в результате старости, болезней, уничтожения хищниками, миграции).
Если по каким-то причинам невозможно определить численность популяции, тогда определяют ее плотность, выражаемую как количество особей на единицу площади (в наземных местообитани-ях) или объема (в водных местообитаниях).
Ареал и численность особей в популяции определяется географическими, физическими и экологическими условиями их обитания. Как и в случае видов, распространение популяций является неравномерным. Поэтому различают «сгущенную» и «островную» формы распределения популяций. В случае «сгущенной» формы распространения популяций их ареал характеризуется наличием особенно плотно заселенных зон. Например, «сгущенное» распределение характерно для популяций берез на русской равнине, где они формируют так называемые высокоплотные на единицу площади чистые березняки наряду с уменьшенным количеством растений в других местах ареала. Напротив, «островное» распределение заключается в очаговом распределении берез в Западной Сибири.
Для популяций характерны различия по возрасту. Например, древесные растения в лесах характеризуются разным возрастом, вследствие чего в перекрестном опылении участвует пыльца разновозрастных растений. Напротив, популяции отдельных видов всегда являются молодыми. Например, популяции дальневосточных лососей в море всегда представлены организмами одного возраста.
Половой состав популяций (соотношение полов) организмов (животных и двудомных растений) разных видов также характеризуется различиями (рис. 210), что отражается на интенсивности их размножения. У тех же организмов, у которых существует партеногенез (ракообразные, насекомые), интенсивность размножения является очень высокой.
Численность популяции представляет собой величину, часто меняющуюся в зависимости от условий обитания, частот рождения, гибели и притока организмов из одной популяции в другую. Иногда разные популяции одного вида объединяются или разделяются на более мелкие.
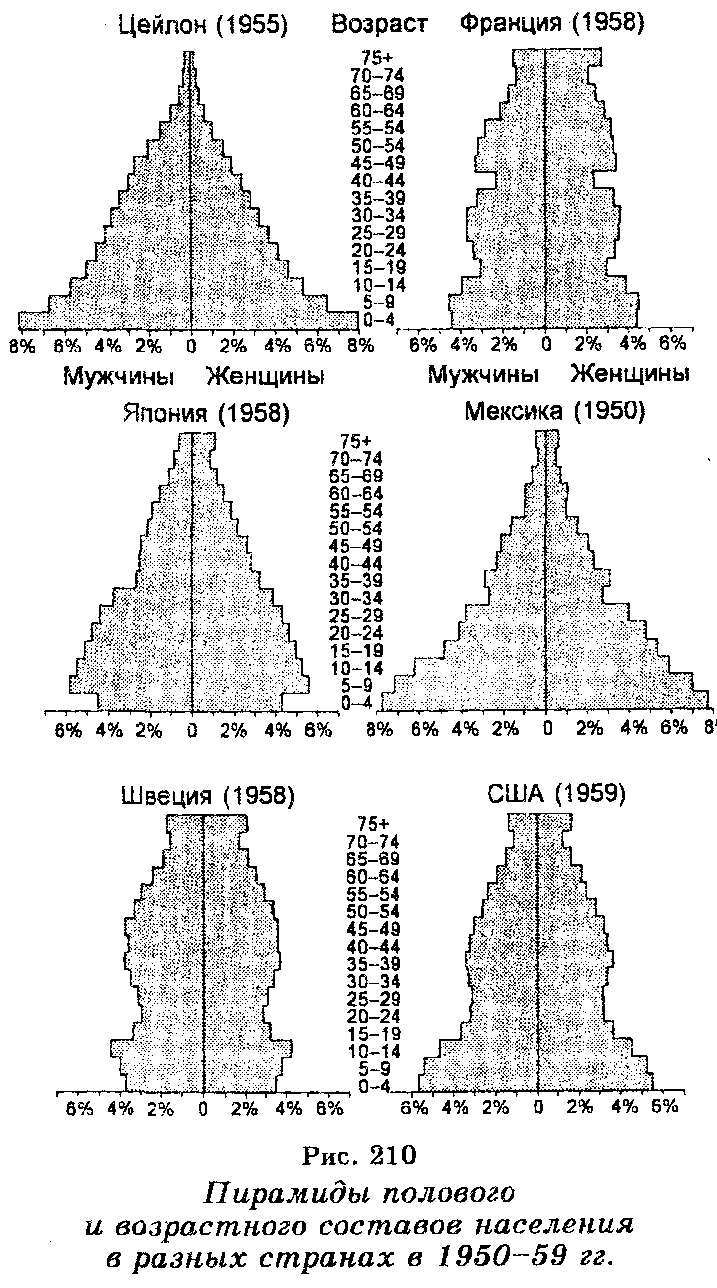 И животные и растения способны очень быстро увеличить свою численность в течение определенного промежутка времени. Эту способность, т. е. их плодовитость, называют биотическим потенциалом вида, который особенно велик, например, у бактерий. Несколько меньшим, но все же очень большим он является у насекомых и некоторых ракообразных, численность которых за год может возрасти примерно в 1030 раз. Что касается млекопитающих, то их биотический потенциал является небольшим. Например, одна пара овец за год в среднем может дать лишь одного ягненка.
И животные и растения способны очень быстро увеличить свою численность в течение определенного промежутка времени. Эту способность, т. е. их плодовитость, называют биотическим потенциалом вида, который особенно велик, например, у бактерий. Несколько меньшим, но все же очень большим он является у насекомых и некоторых ракообразных, численность которых за год может возрасти примерно в 1030 раз. Что касается млекопитающих, то их биотический потенциал является небольшим. Например, одна пара овец за год в среднем может дать лишь одного ягненка.
Если рост популяции происходит в условиях избытка пищи, достаточного места и других благоприятных факторов, то рост численности происходит в геометрической прогрессии или экспоненциально. Классическим примером экспоненциального роста численности являются вспышки численности саранчи (волн жизни). Однако во времени
экспоненциальный рост численности обычно является коротким, после чего он значительно замедляется.
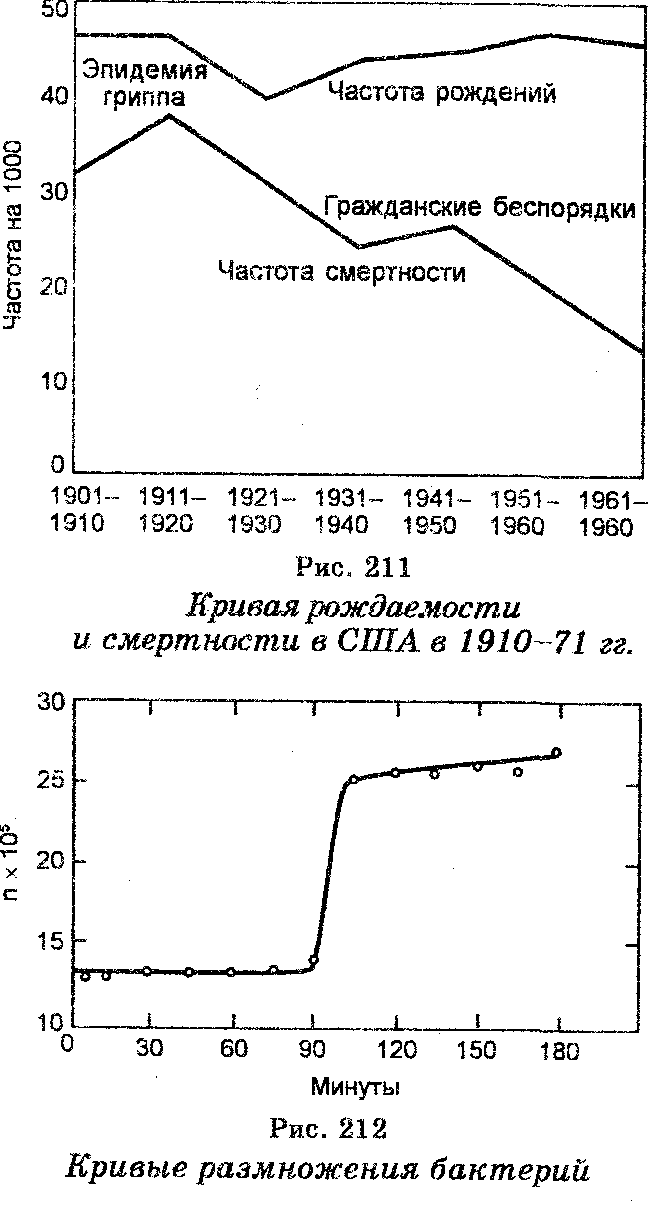
Установлено, что повышение плотности популяции сопровождается уменьшением условий для ее роста и размножения, в результате чего рост численности замедляется (рис. 211). Очень хорошо это положение иллюстрируют также кривые размножения культур бактерий (рис. 212).
 В мире микроорганизмов прирост численности, т. е. плотности культуры зависит от скорости деления клеток. Что же касается многоклеточных организмов, то рост численности зависит от рождаемости и от смертности. По существу, коэффициент рождаемости отражает степень плодовитости.
В мире микроорганизмов прирост численности, т. е. плотности культуры зависит от скорости деления клеток. Что же касается многоклеточных организмов, то рост численности зависит от рождаемости и от смертности. По существу, коэффициент рождаемости отражает степень плодовитости.
Различают абсолютную и удельную рождаемость, причем под первой понимают количество особей, рождающихся в популяции в единицу времени, тогда как под второй понимают количество родившихся особей на определенное число организмов. В случае человека удельную рождаемость выражают коэффициентом рождаемости. Например, если рождается 5,6 детей на 100 жителей, коэффициент рождаемости составит 5,6%.
В противоположность рождаемости смертность определяют как скорость уменьшения численности популяции вследствие гибели отдельных организмов в результате старости, болезней, хищников и т. д.
Изменение численности организмов в замкнутых популяциях зависит от соотношения смертности и рождаемости. При смертности, большей рождаемости, рост численности становится отрицательным. Напротив, при рождаемости, превышающей смертность, рост численности становится положительным, т.е. численность популяции увеличивается.
Численность популяций всегда подвержена колебаниям, частота которых наиболее высокая у насекомых. Например, колебания численности саранчи в годы ее интенсивного размножения (волн жизни) составляют сотни тысяч раз. У млекопитающих колебания численности являются небольшими.
Эффективность действия одних факторов на численность зависит от плотности, действие же других с плотностью не связано. Например, обеспеченность пищей или распространение инфекций зависит от плотности организмов, тогда как губительный характер снежных зим для птиц не зависит от их плотности.
Дата добавления: 2015-12-29; просмотров: 1914;