Стабильность биосферы
Основой самоподдержания жизни на Земле являются биогеохимические круговороты. Процессы созидания органического вещества, аккумулирующего энергию, и противоположные процессы его разложения с высвобождением этой энергии одинаково необходимы для существования жизни. Все доступные для живых организмов химические соединения в биосфере имеют предел. Исчерпаемость пригодных для усвоения химических веществ часто тормозит развитие тех или иных групп организмов в локальных участках суши или океана. По выражению академика В. Р. Вильямса, единственный способ придать конечному свойства бесконечного – это заставить его вращаться по замкнутой кривой. Все химические элементы, используемые в процессах жизнедеятельности организмов, совершают постоянные перемещения, переходя из живых тел в соединения неживой природы и обратно. Возможность многократного использования одних и тех же атомов делает жизнь на Земле практически вечной при условии постоянного притока нужного количества энергии.
Глобальный биогеохимический круговорот вещества на планете представляет собой систему сложно переплетенных циклов отдельных элементов, связанных друг с другом по аналогии с деталями часового механизма (рис. 163). Объединяясь в разных пропорциях в составе сложных органических молекул, синтезируемых в живых телах, атомы различных элементов вне живого вещества имеют свою «судьбу» в атмосфере, гидросфере и литосфере. Круговороты планетарного масштаба создаются из бесчисленных локальных циклических перемещений атомов, движимых жизнедеятельностью организмов в отдельных экосистемах, и тех перемещений, которые вызываются действием ландшафтных и геологических причин (поверхностный и подземный сток, ветровая эрозия, движение морского дна, вулканизм, горообразование и т. п.). Различают поэтому малые и большие круговороты в соответствии с объемом рассматриваемых экосистем: локальные биологические круговороты, биогеохимические циклы материков и океанов и, наконец, общепланетарный биогеохимический круговорот.
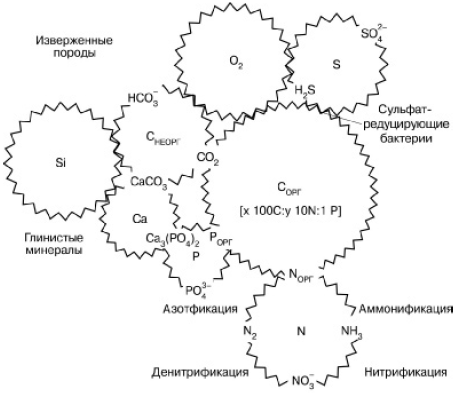
Рис. 163. Сопряжения циклов элементов в биосфере (по Г. А. Заварзину, 1998)
Вместе с тем отличительная черта биологических круговоротов – их неполная замкнутость. Часть химических элементов и их соединений постоянно выпадает из общей циркуляции и скапливается вне организмов, создавая своего рода запасы биогенных веществ. Так были накоплены кислород и азот в атмосфере, горючие ископаемые и другие породы земной коры. В масштабах геологического времени даже небольшое неравновесие в создании и разрушении органических веществ приводит к направленному преобразованию поверхностных слоев Земли.
Запасы биогенных веществ, мало доступные для живых организмов и медленно вовлекающиеся поэтому в биологический круговорот, составляют так называемый резервный фонд того или иного химического элемента в биосфере (например, кальция в карбонатных породах). Другая часть, интенсивно циркулирующая между телами организмов и окружающей средой, называется подвижным, или обменным, фондом. Соотношения между резервным и обменным фондами в биосфере у всех биогенных элементов различны.
Углерод.Углерод как химический элемент является основой жизни. Он может соединяться разными способами со многими другими элементами, образуя простые и сложные органические молекулы, входящие в состав живых клеток. По распространению на планете углерод занимает одиннадцатое место (0,35 % от веса земной коры), но в живом веществе в среднем составляет около 18 или 45 % от сухой биомассы.
В атмосфере углерод входит в состав углекислого газа СО2, в меньшей мере – в состав метана СН4 или следового количества других газообразных соединений. В гидросфере СО2 растворен в воде, и общее его содержание намного превышает атмосферное. Океан служит мощным буфером регуляции СО2 в атмосфере: при повышении в воздухе его концентрации увеличивается поглощение углекислого газа водой. Некоторая часть молекул СО2 реагирует с водой, образуя угольную кислоту, которая затем диссоциирует на ионы НСО3– и СО2-3. Эти ионы реагируют с катионами кальция или магния с выпадением карбонатов в осадок. Подобные реакции лежат в основе буферной системы океана, поддерживающей постоянство рН воды. При подкислении (увеличении концентрации ионов Н+) происходит сдвиг влево в цепи: СО2 воздуха → СО2 воды → Н2СО3 → Са(НСО3)2 → СаСО3. При подщелачивании усиливается выпадение в осадок карбонатов кальция.
Углекислый газ атмосферы и гидросферы представляет собой обменный фонд в круговороте углерода, откуда его черпают наземные растения и водоросли (рис. 164). Фотосинтез лежит в основе всех биологических круговоротов на Земле. Высвобождение фиксированного углерода происходит в ходе дыхательной активности самих фотосинтезирующих организмов и всех гетеротрофов – бактерий, грибов, животных, включающихся в цепи питания за счет живого или мертвого органического вещества.
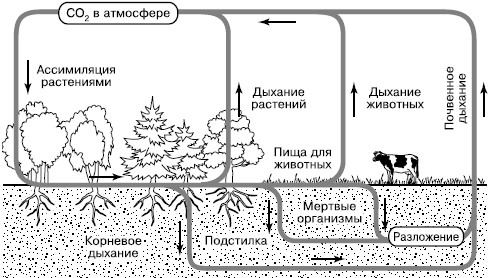
Рис. 164. Круговорот углерода в биосфере (по Б. Болину, 1972)
Особенно активно происходит возврат в атмосферу СО2 из почвы, где сосредоточена деятельность многочисленных групп деструкторов и редуцентов и осуществляется дыхание корневых систем растений. Этот интегральный процесс обозначается как «почвенное дыхание» и вносит существенный вклад в пополнение обменного фонда СО2 в воздухе. Параллельно с процессами минерализации органического вещества в почвах образуется гумус – богатый углеродом сложный и устойчивый молекулярный комплекс. Гумус является носителем почвенного плодородия, поскольку разрушается определенными группами микроорганизмов медленно и постепенно, обеспечивая равномерное питание растений. Гумус почв является одним из важных резервуаров углерода на суше.
В тех условиях, где деятельность деструкторов тормозится факторами внешней среды (например, при возникновении анаэробного режима в почвах и на дне водоемов), органическое вещество, накопленное растительностью, не разлагается, превращаясь со временем в такие породы, как каменный или бурый уголь, торф, сапропели, горючие сланцы и другие, богатые накопленной солнечной энергией. Они пополняют собой резервный фонд углерода, надолго выключаясь из биологического круговорота. Углерод временно депонируется также в живой биомассе, в мертвом опаде, в растворенном органическом веществе океана и т. п. Однако основным резервным фондом углерода на планете являются не живые организмы и не горючие ископаемые, а осадочные породы – известняки и доломиты. Их образование также связано с деятельностью живого вещества. Углерод этих карбонатов надолго захоранивается в недрах Земли и поступает в круговорот лишь в ходе эрозии при обнажении пород в тектонических циклах.
В биологическом круговороте участвуют лишь доли процента углерода от общего его количества на Земле. Углерод атмосферы и гидросферы многократно проходит через живые организмы. Растения суши способны исчерпать его запасы в воздухе за 4–5 лет, запасы в почвенном гумусе – за 300–400 лет. Основной возврат углерода в обменный фонд происходит за счет деятельности живых организмов, и лишь небольшая часть его (тысячные доли процента) компенсируется выделением из недр Земли в составе вулканических газов.
В настоящее время мощным фактором перевода углерода из резервного в обменный фонд биосферы становится добыча и сжигание огромных запасов горючих ископаемых.
По учетам в сети глобального мониторинга это уже приводит к повышению концентрации СО2 в атмосфере, последствия чего для судьбы человеческого общества усиленно обсуждаются и требуют научно обоснованного прогнозирования.
Кислород.С углеродным циклом теснейшим образом сопряжен круговорот кислорода.
Своей уникальной среди планет атмосферой с высоким содержанием свободного кислорода Земля обязана процессу фотосинтеза. Кислород освобождается из молекул воды и является по сути дела побочным продуктом фотосинтетической активности растений. Абиотическим путем кислород возникает в верхних слоях атмосферы за счет фотодиссоциации паров воды, но этот источник составляет лишь тысячные доли процента от поставляемыми фотосинтезом. Между содержанием кислорода в атмосфере и гидросфере существует подвижное равновесие. В воде его примерно в 21 раз меньше на равный объем по сравнению с воздухом.
Выделившийся кислород интенсивно расходуется на процессы дыхания всех аэробных организмов и на окисление разнообразных минеральных соединений. Эти процессы происходят в атмосфере, почве, воде, илах и горных породах. Показано, что значительная часть кислорода, связанного в осадочных породах, имеет фотосинтетическое происхождение. Обменный фонд О2 в атмосфере составляет не более 5 % от общей продукции фотосинтеза. Многие анаэробные бактерии также окисляют органические вещества в процессе анаэробного дыхания, используя для этого сульфаты или нитраты.
Накопление кислорода в атмосфере и гидросфере происходит в геологической истории в результате неполной замкнутости цикла углерода. На полное разложение органического вещества, создаваемого растениями, требуется точно такое же количество кислорода, которое выделилось при фотосинтезе. Захоронение органики в осадочных породах, углях, торфах послужило основой поддержания обменного фонда кислорода в атмосфере. Весь имеющийся в ней кислород проходит полный цикл через живые организмы примерно за 2000 лет.
В настоящее время значительная часть кислорода атмосферы связывается в результате работы транспорта, промышленности и других форм антропогенной деятельности. По расчетам, человечество тратит уже более 10 млрд т свободного кислорода из общего количества в 430–470 млрд т поставляемых процессами фотосинтеза. Если учесть, что в обменный фонд поступает лишь небольшая часть фотосинтетического кислорода, деятельность людей в этом отношении начинает приобретать угрожающие масштабы.
Вода.В ходе фотосинтеза растения используют водород воды в построении органических соединений, освобождая молекулярный кислород. В процессах дыхания всех живых существ, при окислении органических соединений вода образуется вновь. В истории жизни вся свободная вода гидросферы многократно прошла циклы разложения и новообразования в живом веществе планеты.
Кроме биологических циклов на Земле также осуществляется глобальный круговорот воды, движимый энергией Солнца. Вода испаряется с поверхности водоемов и суши и затем вновь поступает на Землю в виде осадков. Над океаном испарение превышает осадки, над сушей – наоборот. Эти различия компенсируются речным стоком. В глобальном круговороте воды растительность суши играет немаловажную роль. Транспирация растений на отдельных участках земной поверхности может составить до 80–90 % выпадающих здесь осадков, а в среднем по всем климатическим поясам – около 30 %. В истории биосферы появление наземной растительности вызвало за счет транспирации перераспределение осадков над морем и сушей и тем самым косвенно повлияло на размеры речного стока и процессы физического и химического выветривания.
Азот.В атмосфере и живом веществе содержится менее 2 % всего азота на Земле, но именно он поддерживает жизнь на планете. Азот входит в состав важнейших органических молекул – ДНК, белков, липопротеидов, АТФ, хлорофилла и других. В растительных тканях его соотношение с углеродом составляет в среднем 1: 30, а в морских водорослях 1: 6. Биологический цикл азота поэтому также тесно связан с углеродным.
Молекулярный азот атмосферы недоступен растениям, которые могут усваивать этот элемент только в виде ионов аммония, нитратов или из почвенных или водных растворов. Поэтому недостаток азота часто является фактором, лимитирующим первичную продукцию. Тем не менее атмосферный азот широко вовлекается в биологический круговорот благодаря деятельности прокариотических организмов (рис. 165). Способностью к фиксации молекулярного азота обладают очень многие прокариоты. В большой мере она развита у фотосинтезирующих сине-зеленых водорослей (цианобактерий). Активно фиксируют азот свободно живущие в почве бактерии рода Azotobacter, а также клубеньковые бактерии Rhizobium, живущие на корнях растений семейства бобовых. При этом может связываться до 400 кг азота на га в год. Отмирая, бактериальные клетки обогащают почву азотными соединениями, доступными для растений. Симбиотические формы снабжают хозяина и за счет прижизненных выделений.
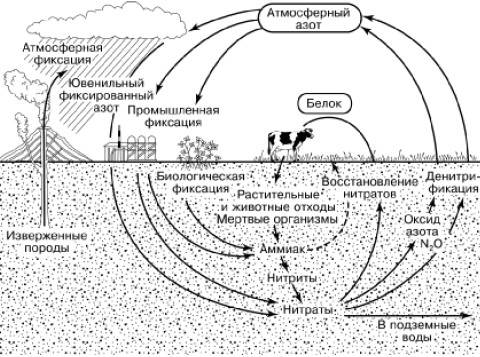
Рис. 165. Круговорот азота в биосфере (по К. Делвичу, 1972)
В круговороте азота принимают большое участие также аммонифицирующие микроорганизмы. Они разлагают белки и другие содержащие азот органические вещества до образования аммиака. В аммонийной форме азот частью вновь поглощается корнями растений, а частью перехватывается нитрифицирующими микроорганизмами. Они используют для себя химическую энергию, окисляя аммиак сначала в нитритную форму (бактерии Nitrosomonas). Окисление нитритов в нитраты производят бактерии рода Nitrobacter. Образовавшиеся нитраты вновь используются растениями в ходе фотосинтеза.
Прямо противоположна по функциям группа микроорганизмов – денитрификаторов.
В анаэробных условиях в почвах или водах они используют кислород нитратов для окисления органических веществ, получая энергию для своей жизнедеятельности. Азот при этом восстанавливается до молекулярного. Азотфиксация и денитрификация в природе приблизительно уравновешены. Цикл азота, таким образом, зависит преимущественно от деятельности бактерий, тогда как растения встраиваются в него, используя промежуточные продукты этого цикла и намного увеличивая масштабы азотной циркуляции в биосфере за счет продуцирования своей биомассы. Объемы микробной фиксации азота составляют до 2,5 т на км2 в год.
Небиологическая фиксация азота и поступление в почвы его окислов и аммиака происходит также с дождевыми осадками при ионизации атмосферы и грозовых разрядах. В среднем это дает около 1 т связанного азота на км2 в год.
Современная промышленность удобрений фиксирует азот атмосферы в размерах, превышающих природную азотфиксацию в целях увеличения продукции сельскохозяйственных растений.
Высокие дозы азотных удобрений приводят, однако, к вымыванию нитратов в грунтовые воды, водоемы и в конечном счете – питьевую воду, а также к избытку их в продуктах питания, что является опасным для человека. Таким же источником загрязнений служат сточные воды с высоким содержанием аммония. На его окисление до нитратов тратится растворенный в воде кислород, что часто бывает губительно для гидробионтов. Таким образом, деятельность человека все сильнее влияет на круговорот азота, в основном в сторону превышения перевода его в связанные формы над процессами возврата в молекулярное состояние.
Фосфор.Этот элемент, необходимый для синтеза многих органических веществ, включая АТФ, ДНК, РНК, усваивается растениями только в виде ионов ортофосфорной кислоты (РО3+4). Он относится к элементам, лимитирующим первичную продукцию и на суше, и особенно в океане, поскольку обменный фонд фосфора в почвах и водах невелик. Круговорот этого элемента в масштабах биосферы незамкнут.
На суше растения черпают из почвы фосфаты, освобожденные редуцентами из разлагающихся органических остатков, но в щелочной или кислой почве растворимость фосфорных соединений резко падает. Основной резервный фонд фосфатов содержится в горных породах, созданных на дне океана в геологическом прошлом. В ходе выщелачивания пород часть этих запасов переходит в почву и в виде взвесей и растворов вымывается в водоемы. В гидросфере фосфаты используются фитопланктоном, переходя по цепям питания в других гидробионтов. Однако в океане большая часть фосфорных соединений захоранивается с остатками животных и растений на больших глубинах и не попадает вновь в фотическую зону, переходя с осадочными породами в большой геологический круговорот. На глубине растворенные фосфаты связываются с кальцием, образуя фосфориты и аппатиты. В биосфере, по сути дела, происходит однонаправленный поток фосфора из горных пород суши в глубины океана, и обменный фонд его в гидросфере очень ограничен. Биологическая продуктивность океана резко повышается в районах, где увеличивается доступность фосфора для фитопланктона – при сносе его речными водами, у побережий морей и в зонах так называемого апвеллинга – восходящих океанических течений, возвращающих фосфорные соединения со дна в освещенные слои воды.
Наземные залежи фосфоритов и аппатитов разрабатываются в качестве удобрений. Снос избыточного фосфора в пресные водоемы является одной из главных причин их «цветения» – бурной эвтрофикации.
Сера.Круговорот серы, необходимой для построения ряда аминокислот, ответственных за трехмерную структуру белков, поддерживается в биосфере широким спектром бактерий. В отдельных звеньях этого цикла участвуют аэробные микроорганизмы, окисляющие серу органических остатков до сульфатов, а также анаэробные сульфатредукторы, восстанавливающие сульфаты до сероводорода, кроме них – разные группы серобактерий, окисляющих сероводород до элементарной серы и далее – сульфатов, тионовые бактерии, переводящие элементарную серу также в соли серной кислоты. Растения усваивают из почвы и воды только ионы SO2-4, поставляемые им деятельностью прокариотов.
Основное накопление серы происходит в океане, куда сульфатные ионы непрерывно поступают с суши с речным стоком. Частично сера возвращается в атмосферу при выделении из вод сероводорода и окисляется здесь до двуокиси, превращаясь в дождевой воде в серную кислоту. Промышленное использование большого количества сульфатов и элементарной серы и сжигание горючих ископаемых поставляют в атмосферу большие объемы диоксида серы. Это вредит растительности, животным, людям и служит источником кислотных дождей, усугубляющих отрицательные эффекты вмешательства человека в круговорот серы.
Как мы видим, круговороты всех биогенных элементов на планете поддерживаются сложным взаимодействием разных частей биосферы. Они формируются деятельностью разных по функциям групп организмов, системой стока и испарения, связывающих океан и сушу, процессами циркуляции вод и воздушных масс, действием сил гравитации, тектоникой плит и другими масштабными геологическими и геофизическими процессами. Биосфера действует как единая сложная система, в которой циркулируют с разной скоростью атомы отдельных элементов. Однако главным двигателем этих круговоротов является живое вещество планеты, все живые организмы, обеспечивающие процессы синтеза, трансформации и разложения органического вещества.
Развитие биосферы
Возраст Земли, определяемый методами изотопной геологии, составляет около 5 млрд лет. Наиболее принятые показатели 4,6–4,7 млрд лет. Приблизительно таков же возраст Солнца и других планет Солнечной системы. По современным представлениям, они образовались из вращающегося газопылевого космического облака. Под влиянием тепла, выделяющегося при гравитационном сжатии и радиоактивном распаде атомов, вещество Земли постепенно дифференцировалось на ядро, мантию и верхние слои. На ранней Земле предполагают активный вулканизм. За счет лав, выплавляющихся из верхней мантии, постепенно сформировалась земная кора, а дегазация лав привела к возникновению первичной атмосферы и жидкой воды на поверхности планеты. В первый миллиард лет существования Земли океан был, по самым грубым оценкам, примерно в 5 раз меньше современного по глубине и объему. Он формировался за счет разрастания и слияния мелких озер на поверхности суши. Ландшафт, подобно современному лунному, представлял вулканические конусы на плоских пространствах. Состав древней атмосферы считают близким к составу газов, выделяющихся из современных вулканов. Они содержат водяной пар (до 70–80 %), углекислый газ (6-19 %), хлор (до 7 %), метан, аммиак, соединения серы и многие другие компоненты. Химический анализ газовых пузырьков в древнейших породах Земли показал полное отсутствие в них свободного кислорода, около 60 % СO2, около 35 % H2S, SO2, NH3, НСl и HF, некоторое количество азота и инертных газов.
В настоящее время имеется уже достаточно много неоспоримых доказательств того, что ранняя атмосфера Земли была бескислородной, аналогично другим планетам Солнечной системы. Ультрафиолетовое излучение Солнца свободно достигало поверхности воды и суши из-за отсутствия озонового экрана. Вулканические газы, растворяясь в воде, переходили в первичный океан, имевший в результате сильно кислую реакцию.
Возникшая на Земле жизнь постепенно изменила эти условия и преобразовала химию верхних оболочек планеты.
Историю Земли делят на три большие отрезка: архей – первые примерно два миллиарда лет ее существования, протерозой – следующие 2 млрд лет и фанерозой, который начался около 570 млн лет назад. Дофанерозойское время называют криптозоем, т. е. эрой скрытой жизни, поскольку древние породы не содержат скелетных отпечатков макроископаемых. Традиционная палеонтологическая летопись начинается лишь с фанерозоя – «эры явной жизни». В более древних слоях присутствуют в основном микроископаемые.
До недавнего времени считалось, что возникновению жизни на Земле предшествовала очень длительная (миллиарды лет) химическая эволюция, включающая спонтанный синтез и полимеризацию органических молекул, объединение их в сложные системы, предшествующие клеткам, постепенное становление обмена веществ и т. п. Возможность и легкость протекания абиогенного синтеза органических мономеров в условиях, моделирующих атмосферу древней Земли, была убедительно доказана еще в 50-х годах во многих лабораториях мира, начиная с известных опытов С. Миллера и Г. Юри. Однако путь от простых органических молекул до простейших живых клеток, обладающих способностью к размножению и аппаратом наследственности, считали очень долгим. К тому же древние породы казались безжизненными. С развитием тонких методов исследования органических молекул, содержащихся в архейских и протерозойских породах, а также остатков микроскопических клеточных структур, это мнение изменилось. Одним из самых удивительных палеонтологических открытий последних десятилетий является регистрация следов жизни даже в самых древних породах земной коры. Следовательно, эволюция от органических соединений до живых клеток протекала в очень сжатые сроки, в самом начале истории Земли. Очень рано появились и фотосинтезирующие организмы. Породы древностью 3,8 млрд лет уже свидетельствуют о наличии на Земле цианобактерий (сине-зеленых водорослей), и следовательно, существовании фотосинтеза и биогенном выделении молекулярного кислорода. На границе архея и протерозоя цианобактерии уже были представлены богатым набором форм, сходных с современными. Наряду с ископаемыми остатками клеток сине-зеленых, в архейских слоях обнаружены следы их масштабной геологической деятельности – породы, сложенные строматолитами. Эти характерные полосчатые и столбчатые окаменелости возникают вследствие функционирования цианобактериальных сообществ, где тесно пространственно объединены фотосинтезирующие сине-зеленые и целый ряд других видов бактерий, деструкторов и хемосинтетиков. Каждая колония, таким образом, представляет отдельную экосистему, в которой сопряжены процессы синтеза и распада органического вещества. Современные строматолиты возникают лишь в крайне экстремальных условиях – в пересоленных или горячих водах, там, где нет более высокоорганизованной жизни (рис. 166).

Рис. 166. Современные строматолиты в осолоненной лагуне (залив Шарк, Западная Австралия)
Таким образом, можно предполагать, что уже в середине архея жизнь на Земле была представлена разнообразными типами прокариот, начинающими влиять на ее геологическую историю. В восстановительной среде кислород, выделяемый цианобактериями, сначала расходовался на окисление разнообразных соединений и не накапливался в свободном виде в атмосфере. При этом аммиак NH3 окислялся до молекулярного азота N2, метан и окись углерода – до СO2, сера и сероводород – до SO2 и SO3. Состав атмосферы постепенно изменялся.
Развитие жизни шло на фоне геологического развития планеты. В архее за счет химического и физического выветривания и эрозии суши началось формирование первых осадочных пород в океане, происходила их гранитизация и сформировались ядра будущих континентальных платформ. По некоторым предположениям, в начале протерозоя они составляли единый континент, названный Мегагеей, и были окружены единым океаном.
Тектоническая активность Земли, как показывает возраст изверженных пород, непостоянна во времени. Короткие эпохи повышенной активности чередуются с более длительными периодами покоя. Такой цикл занимает до 150–500 млн лет. За историю планеты геологи насчитывают 19 тектоно-магматических эпох, четыре из которых приходятся на фанерозой и 15 – на криптозой. В результате шло возрастание неоднородности земной коры. Повышенный вулканизм, горообразовательные процессы или, наоборот, оседание платформ меняли площади мелководий и условия развития жизни. На Земле то ослабевала, то усиливалась климатическая зональность. Следы древних оледенений известны еще с архейской эры.
Считают, что ранняя жизнь имела сначала локальное распространение и могла существовать лишь на небольших глубинах в океане, примерно от 10 до 50 м. Верхние слои, до 10 м, пронизывались губительными ультрафиолетовыми лучами, а ниже 50 м не хватало света для фотосинтеза. Соли древнего океана отличались повышенным содержанием магния по сравнению с кальцием в соответствии с составом пород первичной земной коры. В связи с этим одни из главных осадочных пород архея – магнийсодержащие доломиты. В океане не возникали сульфатные осадки, так как не было анионов окисленной серы. В древних породах много легко окисляющихся, но не окисленных полностью веществ – графита, лазурита, пирита. В архее в результате деятельности анаэробных железобактерий сформировались значительные толщи магнетита, гематита – руд, содержащих недоокисленное двухвалентное железо. Вместе с тем установлено, что кислород, имеющийся в составе этих пород, фотосинтетического происхождения.
Постепенно увеличивающиеся масштабы фотосинтетической активности цианобактерий привели к появлению и накоплению свободного кислорода в окружающей среде. Переход восстановительной атмосферы в окислительную наметился в начале протерозоя, о чем свидетельствуют изменения химического состава земных пород. Железо стало осаждаться в полностью окисленной, трехвалентной форме. Так возникли, например, железистые кварциты-джеспилиты Криворожского бассейна, Курской магнитной аномалии, озеро Верхнее в США и др. Продуктами выветривания на суше становятся красноцветы, содержащие бурое окисное железо. Исчезают пириты, вместо них в океане начинают появляться сульфаты, и морская вода из хлоридной становится хлоридно-карбонатно-сульфатной.
В истории атмосферного кислорода имеют значение несколько его пороговых величин. На Земле, лишенной фотосинтеза, кислород образуется в атмосфере за счет фотодиссоциации молекул воды. Его содержание, по расчетам Г. Юри, не может превышать 0,001 от современного (точка Юри) и автоматически держится на этом уровне. При таком содержании кислорода может существовать только анаэробная жизнь. Появление молекулярного кислорода за счет фотосинтеза сделало возможным для живых клеток процесс дыхания, который представляет намного более эффективный путь высвобождения энергии, чем анаэробное брожение. С этих позиций важна величина 0,01 содержания кислорода от современного уровня – так называемая точка Пастера. Существует целый ряд микроорганизмов, способных переключать свой энергетический обмен с дыхания на брожение и обратно при колебаниях кислорода ниже или выше точки Пастера. Есть предположения, что в раннем и среднем протерозое подобное переключение долгое время служило регулятором содержания О2, так как при повышении его концентрации кислород начинал интенсивно расходоваться на дыхание. Вместе с тем жизнь получила возможность распространиться почти до поверхности водоемов, так как ультрафиолетовые лучи за счет слабого озонового экрана могли теперь проникать на глубины не более метра.
Третье пороговое содержание О2 (точка Беркнера-Маршалла) соответствует 10 % от современного. Оно определяет такую сформированность озонового экрана, при которой потоки жестких ультрафиолетовых солнечных лучей уже не достигают земной поверхности и не препятствуют развитию жизни.
По расчетам 60-х годов, точка Пастера была перейдена менее 1 млрд лет назад, а точка Беркнера-Маршалла – к концу силура, что обеспечило возможность выхода жизни на сушу. Современные исследования заставляют предполагать более раннее развитие событий. По некоторым данным, переход точки Пастера мог произойти уже 2,5 млрд лет назад, а 10-процентное содержание кислорода было достигнуто уже в период 1,8–2,0 млрд лет от современности.
Таким образом, на протяжении более двух миллиардов лет биосфера формировалась исключительно деятельностью прокариотов. Они полностью изменили геохимическую обстановку на Земле: сформировали кислородную атмосферу, очистили ее от токсических вулканических газов, связали и перевели в карбонатные породы огромное количество СO2, изменили солевой состав океана и сформировали громадные месторождения железных руд, фосфоритов и других ископаемых.
Формирование окислительной атмосферы повлекло за собой бурное развитие эукариотической жизни, энергетика которой основана на процессе дыхания (рис. 167). Очевидно, что эукариотная жизнь тесно связана с аэробной средой, подготовленной для нее прокариотами. Первые аэробные организмы могли возникнуть довольно рано в составе цианобактериальных сообществ, которые, по выражению палеонтологов, были своего рода «кислородными оазисами» в анаэробной среде.
Рис. 167. Акритархи из нижнекембрийских отложений (по А. Ю. Розанову, 1986)
В целом выделявшийся ранними фотосинтезирующими организмами кислород был токсичен и смертельно опасен для анаэробных форм жизни. После его накопления в воде и атмосфере анаэробные прокариотные сообщества оказались оттеснены в глубь грунтов, ко дну водоемов, т. е. в локальные местообитания с недостатком О2.
Во второй половине протерозоя в морях появились разные группы одноклеточных водорослей и простейших. Эукариотический фитопланктон усилил масштабы фотосинтеза. В свою очередь, и цианобактерии оставили в это время огромные залежи строматолитов, что свидетельствует об их высокой фотосинтетической активности. В конце протерозоя в морях создавалось уже так много биологической продукции, что на ее основе возникли древние нефтегазоносные залежи.
Последний этап протерозоя, занимающий около 100 млн лет (венд), демонстрирует взрыв многообразия многоклеточных. Возможно, что многоклеточность появилась и раньше, так как пока еще нет ясности в отношении ряда спорных палеонтологических находок, но только в венде возникает огромное разнообразие водных животных и растений достаточно высокой организации. Крупные местонахождения вендской биоты обнаружены в разных регионах мира: Австралии, Южной Африке, Канаде, Сибири, на побережье Белого моря. Среди животных преобладали кишечнополостные и черви, были формы, напоминающие членистоногих, но в целом большинство из них отличалось своеобразным обликом и не встречалось в более поздних слоях (рис. 168). Среди придонных водорослей было много лентовидных слоевищных форм. Отличительная черта всей вендской биоты – бесскелетность. Животные достигали уже крупных размеров, некоторые – до метра, но имели желеобразные студенистые тела, оставившие отпечатки на мягких грунтах. Хорошая и массовая сохранность отпечатков косвенно свидетельствует об отсутствии трупоядов и крупных хищников в вендских биоценозах.


Рис. 168. Отпечатки вендских бесскелетных организмов («фауна Эдиакары») (из Д. Раун, С. Стенли, 1974)
Органическое вещество биогенного происхождения становится постоянным и обязательным компонентом осадочных пород со второй половины протерозоя.
Новая ступень в развитии органического мира – массовое появление у многоклеточных разнообразных наружных и внутренних скелетов. С этого времени датируется фанерозой – «эра явной жизни», поскольку сохранность скелетных остатков в земных слоях позволяет уже более подробно восстанавливать ход биологической эволюции. В фанерозое резко увеличивается воздействие живых организмов на геохимию океана, атмосферы и осадочных пород. Сама возможность появления скелетов была подготовлена развитием жизни. За счет фотосинтеза Мировой океан терял СO2 и обогащался О2, что изменило подвижность целого ряда ионов. В телах организмов в качестве скелетной основы стали откладываться SiO2, СаСО3, MgCO3, Р, Сu, V и другие минеральные компоненты. Присутствие достаточного количества кислорода облегчило и синтез белка коллагена, необходимого для органических скелетов.
Извлекая ряд веществ из водной среды и накапливая их в своих телах, организмы становятся уже не косвенными, а непосредственными создателями многих осадочных пород, захораниваясь на дне водоемов. Накопление карбонатов стало преимущественно биогенным и известковым, поскольку СаСО3 более интенсивно используется для образования скелетов, чем MgCO3. Способность извлекать кальций из воды приобретают очень многие виды. В начале фанерозоя возникли также крупные залежи фосфоритов, созданных ископаемыми с фосфатным скелетом. Химическое осаждение SiO2 также становится биогенным.
В пределах фанерозоя выделяют три эры: палеозой, мезозой и кайнозой, которые, в свою очередь, подразделяют на периоды. Первый период палеозоя – кембрий– характеризуется таким взрывом биологического разнообразия, что он получил название кембрийской революции. Кембрийские породы насыщены многочисленными организмами (рис. 169). За этот период возникли практически все типы ныне существующих животных и целый ряд других, не дошедших до нашего времени. Появились археоциаты и губки, плеченогие, знаменитые трилобиты, разные группы моллюсков, ракушковые рачки, иглокожие и многие другие. Среди простейших возникли радиолярии и фораминиферы. Растения представлены разнообразными водорослями. Роль цианобактерий уменьшилась, так как строматолиты стали мельче и малочисленнее.
В течение ордовика и силура разнообразие организмов в океане нарастало и их геохимические функции становились все более разнообразными. Появились предки позвоночных животных. Рифообразующая роль перешла от строматолитов к коралловым полипам. Основным же событием палеозоя стало завоевание суши растениями и животными.

Рис. 169. Реконструкция биоты раннего кембрия (из А. Ю. Розанова, 1986)
Возможно, что поверхность материков была заселена прокариотами еще в докембрийское время, если учесть выносливость некоторых форм современных бактерий к жесткому излучению. Однако сложные формы жизни смогли освоить сушу только с формированием полноценного озонового экрана. Этот процесс, очевидно, начался в силурийское время, но основным периодом его развития стал девон. Первые наземные растения – сборная группа псилофитов – характеризуются уже целым рядом примитивных анатомо-морфологических приспособлений к обитанию в воздушной среде: возникают проводящие элементы, покровные ткани, устьица и т. п. По другим чертам своего строения псилофиты еще очень похожи на водоросли. Наземная растительность эволюционировала так быстро, что к концу девона в сырых и приводных местообитаниях возникли леса из плауновых, хвощовых и папоротникообразных (рис. 170). Еще раньше на суше появились мхи. Эта споровая растительность могла существовать только во влажных полузатопляемых биотопах и, захораниваясь в анаэробных условиях, оставила залежи нового типа ископаемых – каменных углей.

Рис. 170. Споровые леса девона (из А. Г. Вологдина, 1963)
В морях девона, наряду с бесчелюстными, уже господствовали разные формы рыб. Одна из групп – кистеперые, приобретшие ряд приспособлений к обитанию в мелких, замусоренных отмирающими растениями водоемах, дала начало первым примитивным земноводным. Еще с силура известны первые наземные членистоногие. В девоне уже существовали мелкие почвенные членистоногие, очевидно, потреблявшие гниющую органику. Однако деструкционный процесс на суше был еще недостаточно эффективным, и биологический круговорот – незамкнутым. Массовое захоронение растительной органики и выход ее из системы биологического круговорота повлекли за собой ускоренное накопление O2 в воздухе. По расчетам М. И. Будыко, содержание атмосферного кислорода в начале фанерозоя составляло около трети от современного. В девоне, и особенно в следующем периоде – карбоне, оно достигло современного и даже превзошло его. Карбоновые леса – вершина развития споровой растительности. Они состояли из древовидных плауновых – лепидодендронов и сигиллярий, гигантских хвощевых – каламитов, мощных и разнообразных папоротниковых (рис. 171).

Рис. 171. Споровые леса карбона (из А. Г. Вологдина, 1963)
Только из месторождений Донбасса известно более 400 ископаемых видов. Высокая продукция растений стимулировалась и достаточно большим содержанием СО2 в атмосфере, которое было примерно в 10 раз выше современного. В каменных углях карбона содержится большое количество углерода, изъятого из воздушных запасов СО2 в тот период.
Уже в карбоне возникли растения и животные, способные завоевывать и маловодные пространства суши: первые голосеменные – кордаиты и первые пресмыкающиеся. Воздушную среду освоили первые летающие насекомые. В морях процветали хрящевые и костистые рыбы, головоногие моллюски, кораллы, остракоды и брахиоподы. Конец палеозоя, пермский период, характеризовался резким изменением климатических условий. Интенсивные вулканизм и горообразовательные процессы (завершение Герцинской тектонической эпохи) привели к регрессии моря и высокому стоянию континентов: южного суперконтинента Гондваны и северного – Лавразии. Резко усилилась географическая зональность. В Гондване обнаружены следы обширного оледенения. В Лавразии, в зоне засушливого климата возникают большие площади осадков испарения – гипсов, каменной и калийной соли (месторождения Соликамска), ангидритов, доломитов. В тропических районах, однако, продолжается накопление каменных углей (Кузбасс, Печора, Китай). Споровая растительность приходит в резкий упадок. Масса кислорода в атмосфере сокращается до значений, характерных для начала палеозоя.
На границе палеозойской и мезозойской эр, в конце перми и начале триаса произошло, на фоне смены флор, глубокое обновление морской и наземной фаун. Среди растений господствуют голосеменные – цикадовые, гинкговые и хвойные. Вымирают многие группы земноводных и ранних пресмыкающихся, в морях исчезают трилобиты.
В мезозое начался распад Гондваны на отдельные континенты и расхождение их друг от друга (рис. 172). Середина мезозоя (юра) характеризуется снова расширением мелководий, ровным теплым климатом и ослаблением географической зональности. Юрские леса были по составу значительно разнообразнее карбоновых, менее влаголюбивы и произрастали не только в болотах и по краям водоемов, но и внутри континентов. По долинам и поймам рек они также оставляли залежи каменных углей. Среди позвоночных на суше господствуют рептилии, освоившие также воздушную и вторично водную среду. Возникают различные группы динозавров, птерозавры, ихтиозавры и многие другие формы (рис. 173).
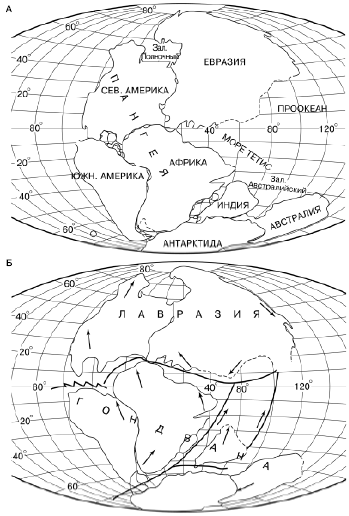
Рис. 172. Положение материков в конце палеозоя и в начале юрского периода (по R. S. Dietz, J. S. Нolden, 1970): стрелки – направление движения; жирными линиями – разломы дна океана

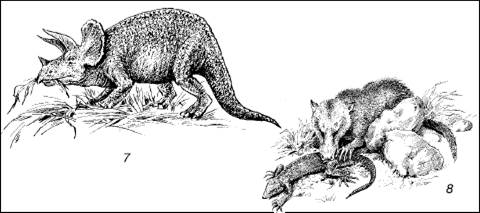
Рис. 173. Рептилии и млекопитающие мезозоя:
1– птеранодон; 2 – рамфоринх; 3 – плезиозавр; 4 – мезозавр; 5 – бронтозавр; 6 – игуанодоны; 7 – трицератопс; 8 – мелкое плотоядное млекопитающее
В мезозое резко сокращается отложение карбонатных пород, одной из причин его считают дальнейшее уменьшение СО2 в атмосфере и океане в связи с расходом на фотосинтез. Меняется и сам характер карбонатных отложений – они представлены в основном биогенным мелом и мергелями с повышенным содержанием Са. В начале мезозоя возникает новая группа одноклеточных водорослей – диатомовые с кремниевыми панцирями и за их счет начинают формироваться тонкие кремниевые илы и новые породы – диатомиты. Их толщи достигают в Мировом океане местами 1600 м при скорости накопления 7-30 см за 1000 лет. Интенсивность фотосинтеза и масштабы захоронения органики очень велики, расходы кислорода на окисление горных пород в межтектонический период незначительны, поэтому к середине мезозоя происходит резкое увеличение массы кислорода в воздухе. По расчетам М. И. Будыко, она превышает современную.
Развитие растительности привело к появлению новой прогрессивной группы – покрытосеменных. Это произошло в меловой период, к концу которого они, быстро распространяясь по всем материкам, значительно потеснили флору голосеменных. Параллельно с цветковыми растениями бурно эволюционируют различные группы насекомых-опылителей и потребителей тканей покрытосеменных. Цветковые растения отличаются ускоренными темпами роста и развития, разнообразием синтезируемых соединений. Будучи независимыми от воды в процессах оплодотворения, они характеризуются тем не менее более высоким потреблением влаги на процессы транспирации, более интенсивным вовлечением в круговорот элементов зольного питания и особенно азота. С появлением растительности покрытосеменных круговорот воды на планете на 80–90 % стал определяться их активностью. Под их влиянием начали формироваться близкие к современным почвы с поверхностным аэробным разложением растительных остатков. Значительно замедлились процессы угленакопления.
В течение всего мелового периода господствовали пресмыкающиеся, многие из которых достигали гигантских размеров. Существовали также зубатые птицы, возникли плацентарные млекопитающие, ведущие свое происхождение еще от примитивных триасовых предков. К концу периода распространились птицы, близкие к современным. В морях процветали костистые рыбы, аммониты и белемниты, фораминиферы.
Конец мелового периода характеризовался началом новой тектонической эпохи и глобальным похолоданием. Смена флор повлекла за собой и смену фаун, усилившуюся в результате влияния глобальных тектонических и климатических процессов. На границе мезозойской и кайнозойской эр произошло одно из наиболее грандиозных вымираний. С лица Земли исчезли динозавры и большинство других рептилий. В морях вымерли аммониты и белемниты, рудисты, ряд планктонных одноклеточных и многие другие группы. Началась интенсивная адаптивная радиация наиболее прогрессивных групп позвоночных – млекопитающих и птиц. В наземных экосистемах большую роль стали играть насекомые.
Наступившая кайнозойская эра характеризовалась возрастанием аэробных условий в биосфере не за счет увеличения массы кислорода, а за счет изменения почвенных режимов. Увеличилась полнота биологических круговоротов. Влажные леса палеогена еще оставили значительные накопления каменных и бурых углей. Одновременно с этим расцвет активной растительности покрытосеменных понизил содержание СО2 в атмосфере до современного уровня, в результате чего снизилась и общая эффективность фотосинтеза. В неогене нарастающий аэробиозис почв и водоемов прекратил процессы образования угля и нефти. В современную эпоху происходит только торфообразование в болотистых почвах.
В течение кайнозоя произошли резкие смены климатов. В результате эволюции покрытосеменных в периоды иссушения в середине эры возникли травянистые растительные формации и новые типы ландшафтов – открытые степи и прерии. В конце усилилась климатическая зональность и наступил ледниковый период с распространением льдов на значительной части Северного и Южного полушарий (рис. 174). Последняя волна ледников отступила всего около 12 тыс. лет назад.

Рис. 174. Ландшафт плейстоцена
В течение всего кайнозоя шла интенсивная эволюция млекопитающих со сменой фаун и вымиранием видов. От одной из групп – приматов около 8–9 млн лет назад обособилась линия, ведущая к человеку. Первые представители рода Homo – Человек – возникли в самом конце кайнозоя, до ледникового периода, около 2,5 млн лет назад (рис. 175). Весь последующий отрезок истории Земли получил название антропогена вследствие особой роли человека в судьбе биосферы.
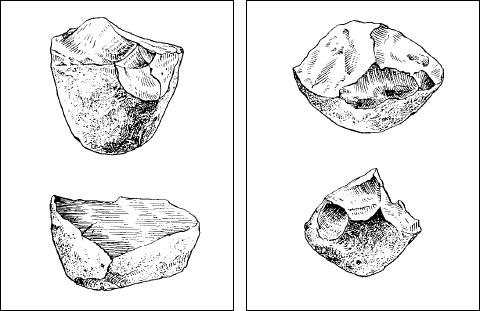
Рис. 175. Древнейшие орудия первых людей (человека умелого)
Дата добавления: 2015-11-26; просмотров: 1126;