Исследование карбонатных пород-коллекторов в шлифах
Изучение карбонатных пород в шлифах является, обязательным в большей степени, чем для других пород, поскольку разнообразные структурные формы этих минералов, в особенности кальцита (органогенный, хемогенный, обломочный), могут быть установлены нередко только микроскопически. Возможность производить на открытых шлифах микрохимические реакции, окрашивание и применение иммерсионного метода еще больше повышает значение этого вида изучения.
Изучение и описание карбонатной породы в шлифах производят по общей схеме: после названия описывают цвет, особенности строения в крупном плане (степень однородности), структуру, текстуру, состав, 262 включения, пористость, вторичные изменения и прочие признаки.
Карбонатная порода, особенно мелко- и микрозернистая, в шлифе часто выглядит серой, что объясняется значительным преломлением карбонатных минералов. Темная тонкорассеянная примесь усиливает серую окраску (рис. 17,18).
| Рис 17. Карбонатная порода при одном николе (х50) | Рис 18. Карбонатная порода в скрещенных николях (х50) |
В шлифе выделяют все структурные разновидности карбонатных пород, включая и самые тонкие — мелко-, микро- и коллоидно- (или коллоидально-) зернистые, а также шламовые и другие, которые макроскопически не различаются и описываются как афанитовые или пелитоморфные. При самых больших увеличениях (до 1000-кратных) становятся различными кокколиты, имеющие размер в десятитысячные доли миллиметра — они нередко являются главной частью писчего мела и мергелей.
Изучение структуры тесно переплетается с изучением состава карбонатных пород и их не всегда легко разделить. Так, определяя и описывая раковинный материал, мы устанавливаем не только структуру породы, но и ее состав как в широком (диалогическом), так и в узком (минералогическом) смысле. Однако все же можно наметить некоторую последовательность при описании структуры и состава.
Почти каждый известняк, как и песчаник, состоит из двух частей — «зерен» (раковин, оолитов, комков и т. д.) и основной массы, являющейся как бы их цементом. Необходимо сначала отметить эту особенность, установить содержание каждой части и затем описать раздельно, начиная с преобладающей, отмечая одновременно и структуру и состав. При описании органогенных компонентов следует отмечать их содержание в породе, степень сохранности (хорошая, гранулированы, перекристаллизованы и т. д.), размер, характер остатков (обломки или целые раковины, окатанность), органогенную форму, минералогический состав и внутреннюю структуру, по которым определятся систематическое положение органических остатков.
Определение может быть самым общим в пределах типа и класса или, если есть возможность, более конкретным, до отряда или семейства. Нередко, однако, приходится давать условные определения или оставлять без названия те или иные остатки организмов. В этом случае они должны быть особенно тщательно описаны и зарисованы, чтобы можно было потом их определить.
При описании оолитов помимо содержания, размера, формы отмечают их строение и состав: наличие центрального зародышевого зерна, его состав, толщина оолитовой части, ее состав, текстура (концентричность, количество и толщина оболочек), структура (зернистая, радиально-лучистая, обычно развивающаяся по зернистой). Необходимо отмечать слуаи раскалывания,окатанность и корродированность оолитов, замещение их другим веществом. Иногда в одном оолите оболочки различаются по составу, примесям, что также не должно ускользать от внимания.
Описание комков, псевдоолитов, сгустков, обломочных карбонатных зерен производят по этой же схеме. Необходимо обращать внимание на их однородность по размеру, форме, структуре, составу и т. д. и стремиться найти признаки, указывающие на их происхождение (остатки организмов, копролиты, оолиты, обломки и т. д.).
Описание основной массы заключается в характеристике ее структуры и состава: размера зерен, степени равно- или разнозернистости, формы зерен, минералогического состава с характеристикой каждого минерала (содержание, форма, размер, диагностические признаки, стадия выделения).
Вместе с карбонатными минералами описывают глинистую примесь. Следует иметь в виду, что ее прежде всего трудно увидеть: более рельефные карбонатные зерна, имеющие яркие цвета интерференции, сильно маскируют глинистые минералы, отличающиеся более низким преломлением и неяркими цветами интерференции. Поэтому глинистая примесь при содержании до 20—25% в шлифе практически не бывает заметна.
Глинистое вещество лучше обнаруживается в тонких участках шлифа, особенно по его краю, где реже мозаика карбонатных зерен. Между этими изометричными зернами становятся видными как в проходящем свете, так и в скрещенных николях удлиненные глинистые чешуйки. Косвенные признаки глинистой примеси — мутный серый цвет породы и микрозернистость. Однако эти признаки могут ввести в заблуждение— так же выглядят и некоторые чистые микрозернистые известняки (например, мел). Более надежный косвенный признак — микрослоистость, обусловливающаяся неодинаковым содержанием глинистого вещества.
Песчано-алеврптовый материал лучше описать отдельно. Его диагностика обычно не представляет трудности. Следует только иметь в виду, что из-за частой корродированности карбонатным веществом обломочные зерна, особенно кварц, можно принять за аутигенные.
Аутигенные минералы, наиболее частые из которых кварц, халцедон, глауконит, фосфаты, пирит, гипс, барит, реже — полевые шпаты, цеолиты, обычно хорошо узнаются. Помимо обычных признаков важно выяснить соотношение их с основными минералами, форму (выполнение пустот, замещение и т. д.) и последовательность выделения.
Диагностика карбонатных минералов:
Главные карбонатные минералы — группа кальцита (кальцит, магнезит, сидерит, доломит, анкерит и др.) —кристаллизуются в тригональной сннгонии, имеют совершенную спайность по ромбоэдру, мало различающуюся по углу (не больше 1—2°), оптически одноосны, отрицательны и обладают сильным двупреломлением, характеризуясь практически одинаковой, перламутровой или белой (розовой) высшего порядка интерференционной окраской и ясным псевдоплеохроизмом в проходящем свете (благодаря изменению рельефа при вращении столика зерна как бы плеохроируют в серых тонах— от бесцветного до серого). Близость ионных радиусов катионов приводит к широкому развитию их изоморфных взаимных замещений и, следовательно, к постепенному изменению не только химического состава, но и оптических свойств, что тоже затрудняет их разделение.
Из другой группы карбонатов — группы арагонита, — кристаллизующихся в ромбической сингонии и являющихся двуосными с небольшим углом оптических осей (практически одноосными), только арагонит принимает заметное участие в составе карбонатных пород. Их оптические свойства (отрицательный знак и очень сильное двупреломление) того же , порядка, что и в группе кальцита. Кристаллографически они псевдогексагональны, т. е. близки к минералам группы кальцита.
Все это свидетельствует о том, что определение карбонатных минералов не может основываться на одном методе, а является комплексным; помимо изучения в образце и в шлифах большое значение имеет иммерсионный метод, химический и термический анализы, а также методы окрашивания.
Остановимся прежде всего на возможностях определения карбонатных минералов в шлифах. Они основываются прежде всего на различиях в кристаллизационной силе карбонатных минералов, а также, отчасти, на различиях цвета, показателей преломления и двупреломления и у арагонита — на его двуосности и субпризматичности. Кристаллизационная сила наименьшая у кальцита, значительно больше у доломита и максимальна у сидерита. В соответствии с этим возрастает степень идиоморфизма и уменьшается размер зерен и разнозернистость структуры.
Известняки часто бывают разнозернисты, нередко в одном шлифе — от крупно- до микрозернистых; форма зерен неправильная, сложная, сильно лапчатая (контур их не только ломаный, но и криволинейный) (рис. 19).
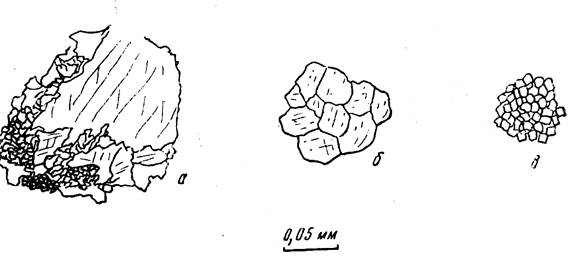
Рис. 19. Структурные различия основных карбонатных пород: а - известняк; б – доломит и анкерит; в - сидерит
Структура доломитов значительно более равномернозер-нистая (см. рис. 19,6), чаще всего просто равномернозернистая, по размеру зерен — от средне- до микрозернистой; реже они бывают крупнозернистыми; форма зерен значительно более простая и правильная, чем у кальцита; она стремится к ромбоэдрической.
Сидерит, если он не сферолитовый, а зернистый, имеет равномернозернистую, микро- или мелкозернистую структуру (см. рис. 19, б) и правильную, субромбоэдрическую форму зерен, которые нередко бывают корродированы минералами гидроокислов железа, часто образующимися при окислении сидерита; окисление идет быстро, и поэтому сидериты, а также анкериты в шлифе всегда имеют розоватую окраску и ассоциируются с железистыми гидроокисными минералами, что служит дополнительным признаком. Естественно, различия структур, обусловленные разной кристаллизационной силой минералов, не универсальны, поскольку они нерезкие. Поэтому на одном этом признаке нельзя основывать определение состава породы. Все же указанный признак важен и позволяет даже в одной породе различать доломит и кальцит: в доломитизированных известняках доломит образует обычно идиоморфные ромбоэдрические кристаллы, потому что кальцит — минерал с меньшей кристаллизационной силой — мешает его росту в меньшей степени, чем доломитовые зерна в чистом доломите. Идиоморфная форма в данном случае не является показателем более раннего па сравнению с кальцитом выделения доломита: нередко он образуется на стадии эпигенеза.
Другой кристаллографической особенностью, помогающей различать кальцит и доломит, является наличие полисинтетических двойников у первого и отсутствие их у доломита (и анкерита, сидерита). Поскольку двойникование проявляется в довольно крупных кристаллах (обычно крупнее 0,1 мм) и чаще образуется при наличии давления (в мраморах кальцит, как правило, полностью сдвойникован), нельзя относить к доломитам породы, карбонат которых не сдвойникован.
К арагониту нередко относят все игольчатые и волокнистые минералы, что часто приводит к ошибкам, поскольку и кальциту присущи эти формы. В шлифах арагонит может быть установлен по его двуосности, хотя из-за малого угла оптических осей для этого необходимы разрезы, в которых по крайней мере одна из оптических осей и острая биссектриса должны быть в поле зрения. Вторым признаком является прямое, по отношению к спайности, погасание арагонита, а у кальцита оно косое, симметричное (рис. 20). Правда, спайность у арагонита выражена не всегда хорошо и часто незаметна, как и в мелких кристаллах кальцита или доломита.
Использование различий преломления и двупреломления карбонатов для их диагностики в шлифах еще менее надежно, однако полезно. Кальцит и доломит (первый чаще, чем второй) дают разрезы с пр меньшим, чем преломление бальзама; у других минералов таких разрезов нет. При одинаковой толщине шлифа кальцит и арагонит имеют более яркую, перламутровую интерференционную окраску, а доломит и другие минералы (из-за их большого двупреломления) — белую или розоватую, в целом более невзрачную интерференционную окраску высшего порядка, хотя по краю шлифа из-за меньшей его толщины и в ряде зерен во всей массе породы вследствие сечения, близкого к перпендикулярному к оптической оси, и доломит имеет перламутровую, т. е. более низкую интерференционную окраску.
Помимо названных различий доломит чаще, чем кальцит, бывает мутным за счет пылеватых непрозрачных включений.
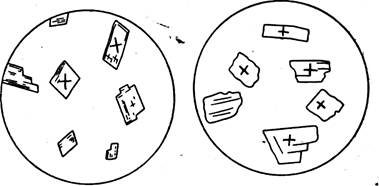
Рис. 20. Форма спайных обломков минералов групп кальцита (К) и арагонита (.4) и направления колебаний света в них ( + ) (Из В. Б. Ттарского
Скелетные породообразующие остатки организмов: основную роль среди них играют остатки животных, меньшую — водоросли, хотя в докембрийские и раннепалеозойские эпохи они, возможно, были основными породообразующими организмами.
Органические остатки определяют по их форме, размеру, внутренней структуре и минеральному составу, в данном случае почти исключительно кальцитовому, реже — арагонитовому.
Структуры:
1. Микрозернистая.
2. Тонкофибровая – устанавливается кальцит по четкому волнистому, нередко правильному крестообразному погасанию можно восстановить их ориентировку, обычно перпендикулярную стенке, в круглых камерах — радиальную.
3. Фибровая, или волокнистая, — у некоторых брахиопод и пелеципод, серпул; волокна становятся видимыми при больших увеличениях, расположение их закономерное, погасание волнистое.
4. Пластинчатая — у многих брахиопод, мшанок, отчасти у пелеципод: пластинки, на поперечном срезе похожие на волокна, ориентированы с разной степенью совершенства поперек стенки, косо или беспорядочно; нередко они гофрированы; погасание чаще всего волнистое, но не четкое.
5. Призматическая — у некоторых брахиопод и многих пелеципод; призмы обычно ориентируются поперек стенки или косо к ней, достигают (у иноцерамов) нескольких миллиметров в толщину и становятся видимыми простым глазом.
6. Крупнокристаллическая — у иглокожих; размеры кристаллов иногда достигают 1 —1,5 см, имеют правильную (круглую, звездчатую и другие) форму и правильную сетчатость, которая облегчает скелет.
Раковина может быть как однослойной, так и многослойной (некоторые брахиоподы, мшанки, кораллы, серпулы, почти все моллюски), образованной слоями с разной структурой.
Остатки животных в известняках представлены камерами фораминифер, спикулами губок, скелетами кораллов, мшанок, члениками-монокристаллами иглокожих, трубочками серпул, раковинами брахиопод и моллюсков, панцирями трилобитов и раковинами остракод и др.
Фораминиферы (рис. 21) —одноклеточные животные микроскопического размера (наиболее мелкие из них имеют размер около 0,01 мм), иногда более крупные, видимые без микроскопа (фузулины — до 3—5 мм, нуммулиты — до 5 см) — обычно имеют внешний известковый многокамерный скелет (реже камеры хитиновые или песчанистые, аглютинированные).
Характерная форма камер и мелкие размеры, способствующие лучшей сохранности, большей частью позволяют безошибочно отличать их от других групп организмов (остракод, серпул, спикул губок, мелких моллюсков и других) и определять с точностью до семейства, отряда, рода и вида.
Дополнительный диагностический признак, на котором в настоящее время ряд палеонтологов начинают строить систематику некоторых групп фораминифер, — структура стенки камер. Им присуща или микрозернистая или тонкофибровая структура. Микрозернистую структуру, благодаря которой стенки часто кажутся темными (см. рис. 21), имеют в основном палеозойские формы (в частности фузулины). Возможно, частично или полностью микрозернистая структура является
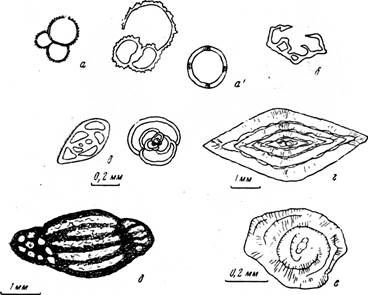
Рис. 21. Раковины фораминифер в шлифах: а, а1 —- глобигерины; б — глоботрунканы; а — милиолиды (мел; около Х50); г — нуммулиты (палеоген; Х10); д — фузулина с микрозернистой стенкой; е — археоднсКуС—с радиально-лучистой стенкой (карбон; Х15 и 80); а' — ник. +, видны крестообразно расположенные четыре участка погасания
вторичной, развившейся по тонкофибровой вследствие грануляции. Фибровая структура присуща большинству фораминифер, включая и некоторые палеозойские формы (Archaeodiscus). Обычно стенка однослойная. Однако иногда развивается два и больше слоев: у некоторых фузулин появляется внутренний слой, отличающийся ячеистостью и наличием выступов внутрь камеры. Многослойность у других является следствием накладывания нескольких оборотов (например, в килевых частях камеры). У третьих имеется внутренний хитиновый слой.
Из других признаков важно отмечать наличие или отсутствие каналов-пор в стенках камер, их размер, форму и расположение; наличие на внешней стороне камер выростов (шипов или игл), которые помогали, парить фораминиферам

Рис. 22. Скелетные остатки кораллов: а — фибровая двуслойная стенка мезозойского (мел) шестилучевого коралла; б— то же — палеозойского одиночного коралла; в — то же, палеозойского колониального коралла; г — гранулированная стенка палеозойского коралла (Х3—5, ник. 1)
в толще воды; толщину стенки, ее соотношение с сечением жилой камеры, чем определяется массивность раковины и условия жизни организма: тонкостенные были планктонными, а формы с массивными раковинами — донными, мелководными (например, нуммулиты). Скелет последних настолько массивен (см. рис. 21, г), что нередко трудно найти жилую камеру: она зажата между массивными оборотами и обычно фи-стончато разделяется столбиками.
Губки имеют внутренний скелет, состоящий из кремневых или известковых спикул, которые чаще всего после смерти организма разобщаются. Известковые спикулы более редки по сравнению с кремневыми, имеют одно-, трех- или четырехлучевую форму, очень тонкий (в отличие от кремневых спикул) осевой канал; погасает спикула одновременно. Размеры спикул чаще микроскопические; они видны при малых увеличениях.
Кораллы имеют внешний арагонитовый скелет, состоящий обычно из двух слоев, которые сложены нормально расположенными фибрами (рис. 22). Фибры соединяются в пучки, последние - в балочки или трабекулы; и те и другие нередко веерообразные, радиально-лучистые, в центре более темные, благодаря чему по оси трабекул проходит темная полоса.

Рис. 23. Скелетные, остатки мшанок: а — поперечное и б — продольное сечения (Крым, даний; Х50, ник. 1)
При фоссилизации арагонит обычно переходит в кальцит, причем фибровое строение в одних случаях сохраняется, в других скелет замещается тонко- или более крупнозернистым кальцитом. Тогда мелкие обломки можно спутать с мшанками, если не видны септы. Кроме того, кораллам свойственна менее правильная и более грубая ячеистость.
Мшанки, широко распространенные с палеозоя до настоящего времени, — колониальные прикрепленные животные с известковым (с примесью MgCO3) внешним скелетом. Форма скелета разнообразная: чаще всего это ветвистые и сетчатые образования, в которых нередко визуально видны ячейки — в них находился организм (зооид). Мшанки, как правило, имеют однослойную тонкопластинчатую стенку, причем пластины, часто изогнутые, ориентированы параллельно стенке (рис. 23). Некоторые из них имеют зернистую структуру, возможно, являющуюся вторичной. В третьих (третичных) В. П. Маслов наблюдал двуслойный скелет: внутренний слой фибровый, с перпендикулярной стенке ориентировкой фибр.
От других ячеистых форм мшанки отличаются довольно правильностью, большей толщиной и структурой стенки; от криноидей — большими размерами и меньшей правильностью ячеек, а также структурой стенки; от водорослей — большими размерами ячеек, часто и структурой стенки; от брахиопод, также имеющих пластинчатое строение стенки, отличаются ячеистостью и более тонкой структурой.
Иглокожие имеют подкожный известковый (с примесью MgC03) скелет, состоящий из крупных (до 1 — 1,5 см) монокристаллических пластин — члеников, обычно правильно перфорированных, и выступающих наружу игл того же строения.. В скрещенных николях они четко отличаются от других остатков погасанием как монокристалла, большими размерами,, округлой, звездчатой и другой правильной формой (рис. 24), тонкой (0,01—0,03 мм) сетчатостью. В ископаемом состоянии членики обычно разобщены; по ним могут быть определены только крупные подразделения.
Наибольшую роль как породообразующие играют членики морских лилий, именно членики стеблей — круглые, овальные, пентагональные или звездчатые в поперечном сечении и четырехугольные — в продольном (вертикальном) (рис. 24), с осевым каналом и правильной или неправильной сетчатостью. Максимально развиты криноидные известняки в девоне, карбоне и юре; есть указания, что они встречаются с докембрия.
Морские ежи, более развитые в мезозое и кайнозое, чем в палеозое, также являются важной породообразующей группой. От члеников морских лилий иглы ежей отличаются удлиненной формой, нередко неодинаковой толщиной, отсутствием канала или, наоборот, широким каналом, более сложной и гармонично построенной сеткой с ясно выраженной на поперечных срезах радиальностью.
Вторичные процессы приводят к стиранию сетчатого строения: ячейки заполняются кальцитом, сначала мелкозернистым, еще выделяющимся при погасании, но потом он приобретает общую с монокристаллом оптическую ориентировку и гаснет одновременно с ним. Часто развивается полисинтетическое двойникование, регенерация и коррозия органогенного кальцита. Однако при внимательном рассмотрении, обычно даже в сильноизмененных породах, можно отличить эхиноидный кальцит от неорганического по реликтам формы, отчасти сетчатости (по загрязнению и т. д.).
Черви иногда имеют известковую (с MgG03) раковину—трубочку, как правило, то более, то менее изогнутую или: спирально закрученную. Они широко распространены в отложениях всех систем, начиная с силура. В меловых и третичных отложениях серпулы участвуют в строении рифов или слагают пласты. Строение стенки, по-видимому, различное
 г
г
Рис. 24. Членики иглокожих: а — поперечное и б — продольное сечения стеблей морских лилий; в — поперечное; г — продольное сечение игл морских ежей (около Х50; ник. 1)
(рис. 25); пластинчатое, зернистое и фибровое радиально-лучистое. Последнее, в частности, присуще верхнемеловым (датским) серпулам Крыма, у которых имеется еще второй микро- (или коллоидально-) зернистый, вероятно, хитиново-известковый слой (см. рис. 25, а).
Брахиоподы (рис. 27) имеют известковую (с небольшой примесью MgC03) или, реже, хитиново-фосфатную (беззамковые оболюсы, лингулы, дисцины) раковину. Строение стенки, вероятно, неодинаковое у разных групп. Преобладают, по-видимому, однослойные раковины, состоящие из
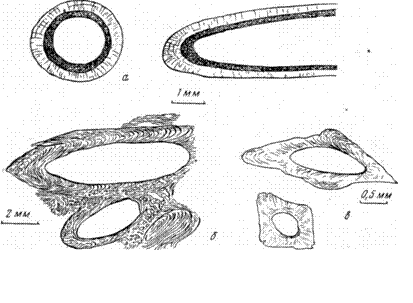 |
Рис 26. Трубочки червей – серпулы; ф – двуслойные, поперечный и диагональный срезы; б – случайное сечение через изогнутую трубочку; в – поперечные и косые сечения
 |
Рис. 27. Обломки раковин брахиопод; а – фосфатная пиритизированная раковина; б – поперечные разрез однослойной раковины; в – косой, поперечный и продольные срезы; г, д – поперечный и тангенциальный срезы пластинчатых раковин; е – поперечный срез трехслойной раковины
тонких (<0;01 мм) или несколько более грубых (0,01— 0,05 мм) пластин, косых или параллельных по отношению к стенке, нередко гофрированных. Реже, например у спириферов, краний, стенка двуслойная и трехслойная: внутренний слой нередко более мощный, а также иногда развивающийся еще третий внешний слой, небольшой мощности, состоят из косо- или перпендикулярно поставленных призм и сходны с призмати ческим слоем пелеципод. Однако у последних он внешний и, кроме того, слабо связан с пластинчатым, благодаря чему легко отделяется и захороняется отдельно. У брахиопод призмы тесно связаны с пластинами, которые как бы продолжаются этими призмами после более или менее резкого изгиба и захороняются обычно вместе. Иглы брахиопод имеют широкий осезон канал и пластинчато-концентрическое строение стенки. В стенках теребратулид, стрингоцефалусов, ортисов и краний имеются канальчики (см. рис. 27).
Фосфатные раковины состоят из перемежающихся тонких, часто выклинивающихся темных (желтых) слоев хитинового вещества и более светлых слоев фосфорнокислого кальция, пересекающихся канальцами. В ископаемых формах хитиновое вещество обычно замещается кальцитом, фосфатом, пиритом или другими минералами.
Моллюски отличаются сложным многослойным и разнообразным строением раковин и только ростры белемнитов наиболее просты: они состоят из радиально расчатый, перламутровый; наиболее внешний — третий (роговой слой)—не сохраняется. У некоторых форм (венериды, кардиды) отсутствует призматический слой, у других — три (пинна и др ) или больше (устрицы, рудисты и лр.) слоев. Состав может быть чисто кальцитовым или чисто арагонитовым, а также одновременно кальцитовым (внешний слой) и арагонитовым. При фоссилизации кальцитовые части макроскопически просвечивают и сохраняют плотную структуру, а арагонитовые утрачивают свою структуру, становятся непрозрачными и похожими на мел.
Призматический слой может быть собственно призматическим, ячеисто-призматическим и пучковидным. Призмы имеют пяти- или шестигранную форму, одинаковую (у иноцерамов) или меняющуюся (у пинны) толщину; иногда достигают значительной длины (у иноцерамов — нескольких сантиметров)
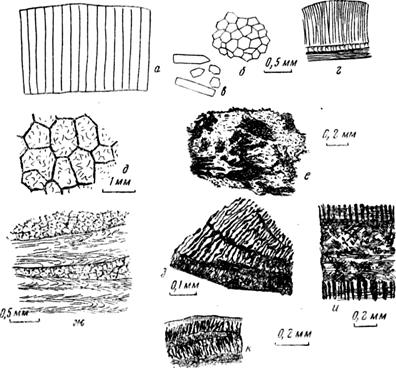
Рис. 28 Обломки раковин моллюсков — двустворок (а—з) и гастропод (и, к); а, б — призматический слой иноцерама на срезе поперечном (а) н параллельном (б) его поверхности; в — отдельные призмы и их обломки (мел, турон, Крым; Х20); г — поперечный срез трехслойной раковины лимы (из Кайе; Х20); д — ячеисто-прпзматический слой рудистов (из Kaiie; Х12); е — пучковидная структура раковины хамы (Х40); ж — чередование ячеисто-призматнческих и пачек волнисто-пластинчатых (складчатых) слоев устрицы (мел, Маастрихт, Крым; Х20); з — кардиум, пластинчатые слон сложной структуры (из Кайе; ХЮО); и —три пластинчатых слоя раковины гастроподы конус (из Кайе; Х45); к — многослойная пластинчатая раковина гастроподы (Х60; везде ник. 1)
и представляют собой монокристаллы. У иноцерамов каждая призма одета тонким слоем органического вещества, что способствует разобщению призм в породе. Ячеисто-призматическая структура характерна для рудистов и некоторых устриц. От собственнопризматической она отличается тем, что призмы обнаруживаются лишь при одном николе (они, как ячеи, по границе очерчены более темным кальцитом), а в скрещенных николях оказываются поликристаллическими, причем границы призм становятся невидимыми. Возможно, это — вторичное явление, развившееся по .призмам арагонита.
Пучковидная структура является сложной и характеризуется наличием пучков пластин. В наиболее простом виде эти пучки напоминают призмы, только не монокристаллические, а агрегатные, листоватые. Пучки, как и пластины, могут выклиниваться, изгибаться, ветвиться, что усложняет рисунок структуры (см. рис. 28).
Пластинчатый слой может быть: 1) листоватым параллельно поверхности раковины, иногда с перпендикулярными линиями раздела, придающими слою решетчатый вид; 2) плетенчатым, решетчатым, когда под разными углами пересекаются две системы пластин; картину усложняют еще линии роста; 3) волнистым, смятым, когда листочки располагаются беспорядочно, сильно изгибаются (грифеи, экзогиры и др.).
Гастроподы имеют арагонитовую (у некоторых имеется внешний кальцптовый слой) трехслойную или многослойную (отчасти, за счет оборотов) раковину, которая в ископаемом состоянии не сохраняет свою внутреннюю структуру. Определяют их по форме раковины и, отчасти, по многослойное и отсутствию структуры слоев. У современных гастропод структура в основном пластинчатая, причем пластины, состоящие из листов, имеют различную ориентировку в разных слоях; нередки слои, сходные с призматическими плетенчатыми слоями пелеципод.
Головоногие (аммониты, гониатиты и наутилоидеи и наутилусы) из-за арагонитового состава раковины также не сохраняют структуру в ископаемом состоянии.
Членистоногие в известняках представлены известковыми панцирями трилобитов и двустворчатыми раковинами остракод микроскопических размеров (доли миллиметра).
Панцирь трилобитов состоит из сегментов, покрывающих различные части тела, включая и конечности. Форма и размер сегментов различные; они часто изогнуты, образуют вздутия или утоняются, а в сечении лентовидны. Структура их тонкофибровая, выявляющаяся волнистым угасанием: волокна ориентированы поперек стенки. При одном николе они обычно однородны, бесцветны, реже сохраняют продольную штриховатость — следы роста, или вертикальные канальчики, которые на тангенциальных срезах кажутся точками.
Растительные известковые остатки многочисленны и разнообразны, особенно в наиболее древних (нижнепалеозойских и докембрнйских) породах. Однако изучены они недостаточно, что объясняется отчасти трудностями их диагностики по скелетным остаткам: форма их разнообразна и меняется даже у одного организма в разных частях в зависимости от условий, т. е. часто нехарактерна, а структура однообразна — все эти водоросли представлены микрозернистым темным кальцитом. Скелет большинства известковых водорослей создается путем отложения микрозернистого кальцита внутри клеток или вне их. Часто такие остатки (корочки, комочки, пленки) плохо отличимы от хемогенного кальцита.
Наибольшее значение для литолога представляют остатки кокколитофорнд, сине-зеленых, зеленых, красных, или багряных, и харовых водорослей.
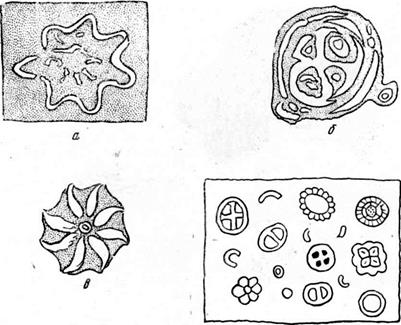
Кокколитофориды — мельчайшие (в сотые доли миллиметра) одноклеточные планктонные жгутиковые водоросли, имеющие известковый панцирь, который состоит из отдельных неделимых — кокколитов. Кокколиты разнообразны по форме (рис. 28); пуговицеобразные, уплощенные часто называют плаколитами, а трубочковидные — рабдолитами. Они известны с силура и живут в современных морях, образуя массовые скопления. В глобигернновом иле, как и в писчем мелу, местами содержание их достигает 70%. В шлифе из-за малых размеров (меньше 0,01 мм) они видны плохо. Желательно изучать их в иммерсионных препаратах. Форма кокколитов лучше видна при одном николе; при скрещенных обнаруживается сферолитовое тонкофибровое строение. В настоящее время их изучают в электронном микроскопе.
г
Рис. 28. Кокколиты: а, б, в — Х2000—3000 (по Коссовской, Шамраю и Лазаревой— из Швецова); г—;<100 (по Швецову)
Сине-зеленые водоросли — низкоорганизованные одноклеточные или многоклеточные нитевидные организмы, известные с докембрия. Ископаемые водоросли делятся на известьотлагающие и сверлящие.
Известьотлагающие сине-зеленые водоросли из-за невозможности сопоставления с современными условно классифицируются по морфологии минеральных остатков. Прежде всего их делят на две группы. Первую составляют онколиты и строматолиты, вторую — собственно сине-зеленые водоросли с ясными следами нитей.
Строматолиты и онколиты являются карбонатными стяжениями, образованными ценозом водорослей, которые почти не оставляют следов для их раздельной диагностики. Обычно они характеризуются неровной (волнистой, бугристой) годичной слоистостью, образованной чередованием более темных (плотных) и более - светлых слоев толщиной в доли миллиметра (рис. 29).
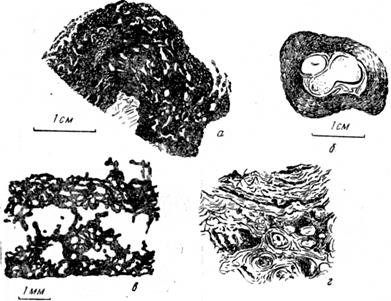
Рис. 29. Сине-зеленые водоросли: а —- онколит (девон; Кузбасс, Х2); б — онколит, образовавшийся вокруг раковины гастроподы (из Маслова); в — строматолит (нижний палеозой Сибири, Х10, ник. 1) (из Маслова); г — строматолит, колония в горизонтальном срезе (натур, величина) (по Маслову)
Строматолиты — прикрепленные и потому нередко протяженные наросты, а онколиты — свободно перекатывающиеся по дну округлые желваки, напоминающие оолиты или пизолиты. Иногда внутри строматолита много пустот, вероятно, от клубков и нитей разных водорослей. Растут строматолиты часто неравномерно, в результате чего участки быстрого роста—столбики — чередуются с впадинами, заполняющимися другим материалом или обломками. Высота столбиков достигает нескольких метров. Наиболее распространенные формы—коллении, конофитоны и другие— часто встречаются в верхнем протерозое и нижнем палеозое.
Собственно сине-зеленые водоросли сохраняют ясные следы нитей, которые обычно остаются в виде канальчиков; заполненных вторичным светлым кальцитом. Стенки этих каналов, как у всех водорослей, темные и состоят из микрозернистого кальцита, отложенного клетками. Для систематики важна величина, форма клеток, наличие поперечных перегородок в каналах. Гирванеллы, относящиеся к группе стелящихся и навивающихся водорослей, представляют собой тонкие (меньше 1 мм) извивающиеся трубочки одинаковой толщины (рис. 30,б). Другая группа объединяет растущие вверх водоросли и поэтому имеющие вид кустика или пучка, например эпифитон, широко распространенный в кембрийских рифах. Его веточки (рис. 30,а) не имеют полости, а состоят из микрозернистого кальцита; возможно, каналы были тонкими и они исчезли в процессе диагенеза.

Рис. 30. Сине-зеленые водоросли. а - эпифотон, б – гирванелла.
Пример описания карбонатных пород
Образец 1. Верхний мел, датский ярус, р. Бодрак, Крым.
Макроскопическое описание. Известняк криноидный, светло-серый, неравнозерннстый, в основном органогенно-обломочнын — мелкодетритовый с более крупными (5 мм) члениками крнноидей (20%), неслоистый, слабо песчанистый, с песчаниковидным и искристым изломом, пористый, средней крепости, бурно вскипает с НС1.
Описание шлифа (рис. 31). Структура крупно-мелкодетритовая, порфировидная: на фоне средне-мелкозернистой (0,5—0,01 мм) д^тритозон основной массы выделяются крупные (2—5 мм) членики криноидей (20%). Текстура беспорядочная.
Состав сложный: помимо органогенного детрита (70—80%) и хемо-генного цементирующего кальцита (10—15%) в породе содержится песчано-алеврнтовая примесь (10—15%).
Органические остатки сложены известковым веществом и представлены иглокожими (40—50%), мшанками (10—15%), фораминнферами (5—10%), а также серпуламн, брахиоподами, пелециподами, водорослями и другими, ближе неопределенными, составляющими вместе не более 5% породы.
Иглокожие представлены в основном члениками криноидей; меньше распространены иглы ежей. Остатки члеников криноидей довольно отчетливо подразделяются на две разновидности — крупные (2—5мм) и мелкие (0,05—0,4 мм). Первые — целые круглые или несколько обломанные правильно-сетчатые (ячейка около 0.01—0,02 мм) монокристаллы с осевым каналом вцентре (0,05—0.1 мм). Вторые — окатанные или угловатые обломки монокристаллических члеников, нередко регенерирован-

Рис. 31. Образец 1. Известняк полидетритовый в основном криноидный (мел, датский ярус, Крым; около Х50. ник. 1): / — криноидей; 2—морские ежи; 3 — мшанки; 4 — форампннферы; 5—серпулы; 6 — брахиоподы; 7 — пелециподы; 8 — водоросли; 9 — кварц; 10 —глауконит
иые по краям чистым хемогенным (несетчатым) кальцитом. В некоторых члениках сетчатость становится плохо различимой. Иглы ежей имеют размер в диаметре 0,05—0,4 мм и характеризуются помимо удлиненной формы более крупной сетчатостью (0,02—0.04 мм), радиальным расположением ячеек, нередко сообщающихся между собой, на поверхности они часто несут многочисленные бугорки.
Мшанки представлены изометричнымн пли удлиненными обломкам
(0,1—2 мм) ячеистой формы. Ячейки по размеру варьируют от 0,05 до 0,2 мм в поперечном сечении и до 1 мм длиной. Стенки их чаще всего тонкие (0,02—0,03 мм), реже — более массивные и сложены тонким пластинчатым кальцитом, ориентированным параллельно стенке. Местами стенка монокристаллична, что свидетельствует о перекристаллизации органогенного кальцита. Ячейки обычно пустые: реже в них развивается микрозернистый кальцит и глауконит.
Фораминиферы представлены целыми мелкими раковинами (0,05—0,2 мм) или их обломками: преобладают спирально-закрученные формы типа кристеллярий, а затем раковины типа текстуллярий. Стенка довольно массивная (0,02—0,03 мм), темно-серая, микрозернистая, очевидно, гранулированная. Реже сохраняется тонкофибровое строение стенки. Камеры пустые и, реже, заполнены мелкозернистым вторичным кальцитом.
Серпулы представлены дугообразными или лентовидными (0,05—2 лии) обломками радиальнолучистого фибрового строения. Нередко сохраняется более темный буроватый внутренний слой (толщиной 0,03—0,07 мм) микрозернистой (или коллоидально-зернистой) структуры и, вероятно, органо-известкового состава. Обломки брахиопод (0,2—2 мм) состоят из тонкопластннчатого, несколько волнистого кальцита, располагающегося параллельно поверхности раковины и, реже, более тонкого призматического слоя, в котором призмы ориентированы перпендикулярно пластинам. Такое двуслойное строение свойственно краниям. К остаткам пелеципод, вероятно, иноцерамов, отнесены довольно редкие удлиненные (0,2 мм) правильные призмы кальцита, а к водорослям — темно-серые равномерно мнкрозернистые обломки, в которых заметна некоторая параллельная и извилистая слоистость Вторичный, хемогенный, кальцит представлен двумя разновидностями: мелкозернистым, заполняющим внутренние полости скелетных остатков, н крупнокристаллическим, иногда регенерационным, заполняющим промежутки между скелетными остатками и песчано-алевритовыми зернами.
Порода сильно пористая (15—20%). Поры изометричные или удлиненные извилистые, размером от 0,01 до 0,3 мм, приурочены как к камерам и ячейкам органических остатков, так и к промежуткам между ними.
Песчано-алевритовая примесь состоит из угловатых и, реже, полуока-танных зерен кварца (около 10%) размером 0,02—0,2 мм, округлых и неправильных зерен свежего глауконита (5%) ярко-зеленого цвета размером 0,1—0,0З мм и единичных зерен полевого шпата.
Из новообразований помимо кальцита присутствует глауконит, заполняющий ячейки мшанок и других органических остатков. Он сходен с зернами глауконита, которые, вероятно, также являются новообразованиями.
Таким образом, изучение в шлифе показало, что известняк является органогенно-обломочным полидетритовым (крупно-средне-мелкозернн-стым), в основном мшанково-криноидным, со значительным содержанием фораминифер, песчанисто-алевритистым (10—15%), сильно пористым, с новообразованиями кальцита и глауконита.
| <== предыдущая лекция | | | следующая лекция ==> |
| Качественный эмиссионный спектральный анализ | | | Исследование обломочных пород-коллекторов в шлифах |
Дата добавления: 2015-11-04; просмотров: 8800;