Окислительно-восстановительный потенциал промежуточных переносчиков и изменение свободной энергии при переносе электронов в дыхательной цепи
| Промежуточные переносчики | Величина окислительно-восстановительного потенциала (Е) | Разница окислительно-восстановительного потенциала (∆Е) | Величина свободной энергии (ккал) | Пункты фосфори-лирования |
| НАД | -0,32 | |||
| ФАД | -0,05 | 0,27 | 12,2 | + |
| Цитохром b | +0,04 | 0,09 | 4,05 | |
| Цитохром с | +0,26 | 0,22 | 9,90 | + |
| Цитохром аа3 | +0,29 | 0,03 | 1,35 | |
| Кислород | +0,82 | 0,53 | 23,80 | + |
Наибольшие значения разности окислительно-восстановительных потенциалов наблюдаются между следующими последовательно расположенными в цепи переносчиками: НАД ð ФАД; цитохром b ð . цитохром с; цитохром аа3 ð кислород. Именно на этих трех этапах переноса происходит наибольшее изменение свободной энергии, за счет которой осуществляется ресинтез трех молекул АТФ, по одной на каждом из указанных участков.
Термодинамические расчеты позволяют оценить эффективность процессов биологического окисления, сопряженного с ресинтезом АТФ. Известно, что при переносе пары электронов от субстрата окисления на кислород освобождается 52,6 ккал/моль энергии (218 кдж). При этом происходит образование 3 молекул АТФ. На ресинтез каждой молекулы АТФ из АДФ и Н3РО4 затрачивается не менее 9 ккал/моль. То есть, на ресинтез АТФ (полезная энергия) в дыхательной цепи затрачивается не менее 27 ккал. Отсюда вытекает, что эффективность процесса составляет 27:52,6.×100% = 51,3%. Это значительно выше, чем эффективность работы различных преобразователей энергии, созданных руками человека.
Исключение составляют субстраты, окисление которых осуществляют дегидрогеназы, содержащие ФАД в качестве кофермента. К числу таких субстратов относятся сукцинат (янтарная кислота), ацил КоА. В процессе переноса водорода с этих субстратов на кислород отсутствует стадия НАД → ФАД, экзергонический эффект которой обеспечивает ресинтез одной молекулы АТФ. Энергетический эффект аэробного окисления таких субстратов составляет не три, а две молекулы АТФ.
Есть основания считать, что фиксация энергии в макроэргических связях АТФ только один из возможных путей использования энергии, освобождающейся при переносе электронов. Кроме того, эта энергия может использоваться непосредственно (без предварительного синтеза АТФ) для обеспечения других эндергонических функций митохондрий: поддержание ионных градиентов и активный транспорт веществ через митохондриальную мембрану, различные биосинтетические процессы и т.п. Конкретные данные о затратах энергии по этим направления деятельности митохондрий пока отсутствуют, поэтому определение итоговой эффективности энергетических процессов в митохондриях не представляется возможным.
Существуют косвенные данные, свидетельствующие о том, что у спортсменов высокой квалификации, специализирующихся в т.н. аэробных видах спорта (бег на длинные дистанции, лыжные и велосипедные гонки и др.), в период высокой спортивной формы эффективность процессов аэробного окисления может повышаться: каждая пара протонов и электронов, проходя по дыхательной цепи, может обеспечить ресинтез не 3, а 4 молекул АТФ. Ресинтез четвертой молекулы АТФ может происходить на этапе переноса электронов от цитохромов аа3 на кислород, где освобождается наибольшее количество энергии. Однако прямых подтверждений этого пока нет.
Ферментные системы, осуществляющие отщепление, транспорт и присоединение протонов и электронов к кислороду, а также сопряжение этих процессов с ресинтезом АТФ, локализованы на кристах - складках внутренней поверхности митохондрий - внутриклеточных образованиях, важнейшей функцией которых является обеспечение аэробного ресинтеза АТФ (см. главу «Общие закономерности обмена веществ»).
Непосредственный механизм фосфорилирования АДФ и образования АТФ достаточно сложен и еще не полностью раскрыт. Существует несколько теорий, объясняющих механизм этого сопряжения. Наибольшее признание (и наиболее полное экспериментальное подтверждение) получила т.н. хемиосмотическая теория, разработанная английским биохимиком П. Митчеллом еще в шестидесятых годах прошлого столетия.
Согласно этой теории, главным сопрягающим фактором экзергонического процесса переноса электронов по дыхательной цепи с эндергоническим процессом ресинтеза АТФ является так называемый протонный насос, действующий на внутренней мембране митохондрий (на кристах), в обычных условиях не проницаемой для ионов Н+. Энергия, освобождающаяся при переносе электронов по дыхательной цепи, используется для энергозависимой перекачки протонов (Н+) через внутреннюю мембрану митохондрий из матрикса в межмембранное пространство и создания протонного градиента - разности концентраций протонов на внутренней и наружной стороне внутренней (сопрягающей) мембраны митохондрий. В результате возникает разность потенциалов (электрохимический потенциал): наружная сторона сопрягающей мембраны заряжена положительно, а внутренняя – отрицательно. Величина электрохимического потенциала установлена экспериментальным путем. Она составляет 0,25В, т.е. вполне достаточна для обеспечения ресинтеза АТФ.
Возникновение протонного градиента стимулирует перемещение ионов водорода в обратном направлении по градиенту концентрации. Однако митохондриальная мембрана непроницаема для протонов. Поэтому это перемещение происходит по специальным каналам, в образовании которых принимает участие один из двух белков, образующих фермент Н+-АТФ-синтетазу.
При этом энергия протонного градиента используется на активацию фосфорной кислоты и ее включение в ресинтез АТФ из АДФ, катализируемый ферментом Н+-АТФ-синтетазой. Синтез АТФ происходит в активном центре АТФ-синтетазы, после чего АТФ перемещается в матрикс митохондрий.
Транспорт АТФ из матрикса митохондрий в межмембранное пространство обеспечивает специфический белок – АТФ/АДФ-транслоказа, локализованный во внутренней мембране митохондрий. Он имеет два центра связывания: со стороны матрикса для АТФ, в межмембранном пространстве – для АДФ. При возникновении определенных условий транслоказа переносит АДФ из межмембранного пространства в матрикс, а АТФ в обратном направлении.
АТФ не может переходить через наружную мембрану митохондрий. Для транспорта макроэргического фосфата в цитоплазму используется креатинфосфатный шунт, механизм которого представлен на рис. 10. 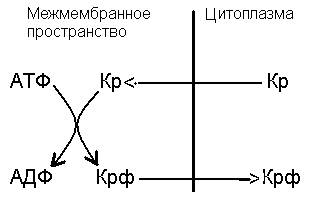
Рис. 10. Механизм переноса макроэргического фосфата через наружную мембрану митохондрий. Кр – креатин, Крф – креатинфосфат.
В цитоплазме креатинфосфат (Крф) вступает в реакцию ресинтеза АТФ, протекающую по уравнению:
Крф + АДФ → Кр + АТФ
Поскольку ресинтез АТФ требует постоянного притока неорганического фосфата, во внутренней мембране митохондрий имеется специальная транспортная система, обеспечивающая перенос неорганического фосфата в матрикс, сопряженая с переносом Н+.
В условиях живого организма при постоянных тратах АТФ и непрерывно идущих процессах биологического окисления и деятельности электронпереносящей цепи устанавливается круговое движение протонов через сопрягающую мембрану, обеспечивающее непрерывный ресинтез АТФ. Скорость этих превращений может существенно меняться в зависимости от потребности в энергии (скорости расходования АТФ).
Дата добавления: 2015-10-29; просмотров: 2828;