Медиаторы и физиология синапсов
В 1897 году Ч.Шеррингтон ввел понятие «синапс» (от греч. «смыкать», «соединять») для обозначения структуры, обеспечивающей передачу возбуждения с нервного волокна на мышечное или с одного нервного волокна на другое. Это место функционального контакта.
Различают периферические и центральные синапсы.
Периферические находятся на границе между нервными волокнами и рабочими органами.
Центральные – между нервными клетками и их структурами. Обычно встречаются синапсы между концевыми веточками аксона одного нейрона и дендритами (аксо-дендритные синапсы) или телом (аксоматические синапсы) другого нейрона.
В зависимости от природы проходящих по синапсам сигналов различают два типа синапсов – электрические и химические. Факты в пользу химической теории передачи нервных импульсов стала накапливаться с 1878 г., в 1906 году Д.Ленгли доказал свою правоту, изучая переход импульса с нервного волокна на мышечное.
Химические синапсы – наиболее распространенный тип синапса у позвоночных. Типичный пример – нервно-мышечный синапс, который существует между окончаниями двигательного нейрона и поверхностью мышечного волокна, это соединение, сходное с межнейронными синапсами в функциональном отношении, но отличающееся по строению. Химический синапс – луковицеобразное утолщение нервных окончаний, названное синаптическими бляшками. Нервно-мышечный синапс состоит из трех основных элементов: пресинаптической мембраны, постсинаптической мембраны и синаптического пространства (Рис.8).
Пресинаптической мембраной служит мембрана нервных веточек – окончаний аксона. Постсинаптическая мембрана – мембрана мышечного волокна. Между ними – синаптическое пространство (щель) шириной примерно 50 нм.
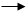
 Как только в нервное окончание проходит нервный импульс, в синаптических пузырьках начинает выделяться медиатор ацетилхолин, пузырьки подходят к мембране, медиатор выходит из них (экзоцитоз) и попадает в синаптическую щель (в каждом пузырьке около 3000 молекул ацетилхолина). Этот процесс называется электросекреторным сопряжением. Медиатор – вещество, с помощью которого нервный сигнал передается через синапс. Ацетилхолин – аммонийное производное, выделенное в 1920 г. Отто Леви из окончаний парасимпатических нейронов блуждающего нерва сердца лягушки. На постсинаптической мембране находится особый белок, образующийся в теле клетки на рибосомах - рецептор (холинорецептор), который чувствителен к ацетилхолину. Холинорецептор соединяется с ацетилхолином, структура постсинаптической мембраны изменяется, повышается ее проницаемость для ионов натрия. Проникая из щели в мышечное волокно, ионы натрия вызывают деполяризацию постсинаптической мембраны, возникает потенциал концевой пластинки (только в нервно-мышечном синапсе). При достижении деполяризации определенного уровня (около 40 мВ), между деполяризованным участком постсинаптической мембраны и соседними внесинаптическими участками с прежним потенциалом, возникает движение кругового электрического тока. Если его сила пороговая – возникает распространяющийся потенциал действия. Ацетилхолинэстераза расщепляет ацетилхолин, в результате гидролиза образуется холин, он всасывается обратно, в синаптическую бляшку, превращается в ацетилхолин, который хранится в пузырьках. Исходное состояние синапса восстанавливается.
Как только в нервное окончание проходит нервный импульс, в синаптических пузырьках начинает выделяться медиатор ацетилхолин, пузырьки подходят к мембране, медиатор выходит из них (экзоцитоз) и попадает в синаптическую щель (в каждом пузырьке около 3000 молекул ацетилхолина). Этот процесс называется электросекреторным сопряжением. Медиатор – вещество, с помощью которого нервный сигнал передается через синапс. Ацетилхолин – аммонийное производное, выделенное в 1920 г. Отто Леви из окончаний парасимпатических нейронов блуждающего нерва сердца лягушки. На постсинаптической мембране находится особый белок, образующийся в теле клетки на рибосомах - рецептор (холинорецептор), который чувствителен к ацетилхолину. Холинорецептор соединяется с ацетилхолином, структура постсинаптической мембраны изменяется, повышается ее проницаемость для ионов натрия. Проникая из щели в мышечное волокно, ионы натрия вызывают деполяризацию постсинаптической мембраны, возникает потенциал концевой пластинки (только в нервно-мышечном синапсе). При достижении деполяризации определенного уровня (около 40 мВ), между деполяризованным участком постсинаптической мембраны и соседними внесинаптическими участками с прежним потенциалом, возникает движение кругового электрического тока. Если его сила пороговая – возникает распространяющийся потенциал действия. Ацетилхолинэстераза расщепляет ацетилхолин, в результате гидролиза образуется холин, он всасывается обратно, в синаптическую бляшку, превращается в ацетилхолин, который хранится в пузырьках. Исходное состояние синапса восстанавливается.
Существуют определенные различия в строении и функционировании возбуждающего и тормозного синапсов:
1. Синаптическая щель тормозного синапса уже, она составляет 20 нм (у возбуждающего - 30 нм).
2. Тормозные синапсы имеют более толстую и плотную постсинаптическую мембрану.
3. Пресинаптическая мембрана тормозного синапса содержит меньше пузырьков медиатора.
4. Медиатором в тормозных синапсах являются глицин, гамма - аминомасляная кислота (ГАМК), иногда - ацетилхолин.
Для включения в работу тормозного синапса опять нужен импульс возбуждения. При достижении импульсом пресинаптической мембраны в синаптическую щель выбрасывается квант тормозного медиатора. Действуя как химический раздражитель на мембранные рецепторы, тормозной медиатор изменяет проницаемость мембраны незначительно (в основном для ионов K+ и Cl-). Поскольку ионов K+ больше внутри клетки, а ионов Cl- в межклеточном пространстве, они начнут двигаться в противоположных направлениях: K+ - наружу, Cl- - внутрь. В результате этого увеличивается поляризованность мембраны (мембранный потенциал). При этом регистрируется положительное колебание с амплитудой около 5 mV - тормозной постсинаптический потенциал (ТПСП). Его продолжительность составляет (как и ВПСП) всего несколько миллисекунд. Но в течение этого времени никакое возбуждение на данном конкретном участке постсинаптической мембраны невозможно. В этом и заключается механизм действия тормозного синапса.
Дополнительно к вышеизложенному, необходимо отметить, что в некоторых нейронах головного мозга постсинаптические потенциалы могут иметь значительно большую длительность: ВПСП - до 80 миллисекунд, ТПСП - до 100 миллисекунд.
Мозг пользуется несколькими медиаторами. Норадреналин, дофамин, серотонин вызывают возбуждение, глицин и гамма - аминомасляная кислота (ГАМК) - торможение. Ацетилхолин - универсальный медиатор. Он может вызывать возбуждение и торможение. Каждый синапс для своей работы использует только один медиатор.
Доля электрических синапсов (эфапсов) в ЦНС высокоорганизованных животных незначительна. Передача возбуждения через эти синапсы называется эфаптической. Пресинаптическая мембрана в таких синапсах сообщается с постсинаптической посредством миниатюрных мостиков, т.е. мембраны не разделены щелью. Потенциал действия, достигнув пресинаптической мембраны, преобразуется в градуальный ток, «стекающий» на постсинаптическую мембрану, где вновь возрождается в форме новой волны возбуждения. Таким путем возбуждение проводится через синапс почти без задержки.
Особенностью эфапсов является также их способность передавать возбуждение в двух противоположных направлениях, что принципиально невозможно в химических синапсах.
В мозговой ткани встречаются и смешанные синапсы. Такой синапс частично может проводить возбуждение как эфапс, а частично - с помощью медиатора.
Полагают, что некоторые лекарственные вещества, используемые при депрессиях и тревожности, воздействуют на химическую передачу в синапсах (Таблица 2).
Многие транквилизаторы и седативные средства (имипрамин, резерпин, др.) производят лечебный эффект, взаимодействуя с медиаторами, их рецепторами или отдельными ферментами. Галлюциногены типа диэтиламида лизерговой кислоты и москалина воспроизводят действие каких-то природных медиаторов мозга или подавляют действие других медиаторов. Сходный эффект с героином и морфием вызывают эндорфины, способные взаимодействовать с опиатными рецепторами. Из эндорфинов лучше всего изучена группа пептидов – энкефалинов (мет-энкефалин, β- эндорфин и др.)
Таблица 2
Дата добавления: 2015-10-13; просмотров: 3621;