Механизмы мембранного транспорта веществ
Различают несколько способов переноса веществ через мембрану:
Простая диффузия – это перенос небольших нейтральных молекул по градиенту концентрации без затрат энергии и переносчиков. Легче всего проходят простой диффузией через липидную мембрану малые неполярные молекулы, такие как О2, стероиды, тиреоидные гормоны. Малые полярные незаряженные молекулы – СО2, NH3, H2O, этанол и мочевина – также диффундируют с достаточной скоростью. Диффузия глицерола идет значительно медленнее, а глюкоза практически не способна самостоятельно пройти через мембрану. Для всех заряженных молекул, независимо от размера, липидная мембрана не проницаема.
Облегченная диффузия – перенос вещества по градиенту концентрации без затрат энергии, но с переносчиком. Характерна для водорастворимых веществ. Облегченная диффузия отличается от простой большей скоростью переноса и способностью к насыщению. Различают две разновидности облегченной диффузии:
а) транспорт по специальным каналам, образованным в трансмебранных белках (например, катионселективные каналы);
б) с помощью белков-транслоказ, которые взаимодействуют со специфическим лигандом, обеспечивают его диффузию по градиенту концентрации (пинг-понг) (перенос глюкозы в эритроциты с помощью белка-переносчика ГЛЮТ-1).
Кинетически перенос веществ облегченной диффузией напоминает ферментативную реакцию. Для транслоказ существует насыщающая концентрация лиганда, при которой все центры связывания белка с лигандом заняты, и белки работают с максимальной скоростью. Поэтому скорость транспорта веществ облегченной диффузией зависит не только от градиента концентраций переносимого вещества, но и от количества беков-переносчиков в мембране.
Простая и облегченная диффузия относится к пассивному транспорту, так как происходит без затраты энергии.
Активный транспорт – транспорт вещества против градиента концентрации (незаряженные частицы) или электрохимического градиента (для заряженных частиц), требующий затрат энергии, чаще всего АТФ. Выделяют два вида его: первично активный транспорт использует энергию АТФ или окислительно-восстановительного потенциала и осуществляется с помощью транспортных АТФ-аз. Наиболее распространены в плазматической мембране клеток человека Na+,K+- АТФ-аза, Са2+-АТФ-аза, Н+-АТФ-аза.
При вторично активном транспорте используется градиент ионов, созданный на мембране за счет работы системы первично активного транспорта (всасывание глюкозы клетками кишечника и реабсорбция из первичной мочи глюкозы и аминокислот клетками почек, осуществляемые при движении ионов Na+ по градиенту концентрации).
Перенос через мембрану макромолекул. Транспортные белки обеспечивают перенос через клеточную мембрану полярных молекул небольшого размера, но они не могут транспортировать макромолекулы, например белки, нуклеиновые кислоты, полисахариды или отдельные частицы. Механизмы, с помощью которых клетки могут усваивать такие вещества или удалять их из клетки, отличаются от механизмов транспорта ионов и полярных соединений.
А) Перенос вещества из среды в клетку вместе с частью плазматической мембраны называют эндоцитоз. Путем эндоцитоза (фагоцитоза) клетки могут поглощать большие частицы, такие как вирусы, бактерии или фрагменты клеток. Поглощение жидкости и растворенных в ней веществ с помощью небольших пузырьков называют пиноцитозом.
Б) Экзоцитоз. Макромолекулы, например белки плазмы крови, пептидные гормоны, пищеварительные ферменты синтезируются в клетках и затем секретируются в межклеточное пространство или кровь. Но мембрана не проницаема для таких макромолекул или комплексов, их секреция происходит путем экзоцитоза. В организме имеются как регулируемый так и не регулируемый пути экзоцитоза. Нерегулируемая секреция характеризуется непрерывным синтезом секретируемых белков. Примером может служить синтез и секреция коллагена фибробластами для формирования межклеточного матрикса.
Для регулируемой секреции характерны хранение приготовленных на экспорт молекул в транспортных пузырьках. С помощью регулируемой секреции происходят выделение пищеварительных ферментов, а также секреция гормонов и нейромедиаторов.
Глава 10
ЭНЕРГЕТИЧЕСКИЙ ОБМЕН. БИОЛОГИЧЕСКОЕ ОКИСЛЕНИЕ
Живые организмы с точки зрения термодинамики – открытые системы. Между системой и окружающей средой возможен обмен энергии, который происходит в соответствии с законами термодинамики. Каждое органическое соединение, поступающее в организм, обладает определенным запасом энергии (Е). Часть этой энергии может быть использована для совершения полезной работы. Такую энергию называют свободной энергией (G). Направление химической реакции определяется значением DG. Если эта величина отрицательна, то реакция протекает самопроизвольно. Такие реакции называются экзергоническими. Если DG положительно, то реакция будет протекать только при поступлении свободной энергии извне – это эндергонические реакции. В биологических системах термодинамически невыгодные эндергонические реакции могут протекать лишь за счет энергии экзергонических реакций. Такие реакции называют энергетически сопряженными.
Важнейшей функцией многих биологических мембран служит превращение одной формы энергии в другую. Мембраны, обладающие такими функциями, называются энергопреобразующими. Любая мембрана, выполняющая энергетическую функцию, способна к превращению химической энергии окисляемых субстратов или АТФ в электрическую энергию, а именно в трансмембранную разность электрических потенциалов (DY) или в энергию разности концентраций веществ, содержащихся в разделенных мембраной растворах, и наоборот. Среди энергопреобразующих мембран, имеющих наибольшее значение, можно назвать внутреннюю мембрану митохондрий, внешнюю цитоплазматическую мембрану, мембраны лизосом и комплекса Гольджи, саркоплазматический ретикулум. Наружная мембрана митохондрий и ядерная мембрана не может превращать одну форму энергии в другую.
Преобразование энергии в живой клетке описывается следующей общей схемой:
Энергетические ресурсы ® DmI ® работа
где DmI – трансмембранная разность электрохимических потенциалов иона I. Следовательно, процессы утилизации энергии и совершения за счет нее работы оказываются сопряжены через образование и использование DmI. Поэтому данный ион может быть назван сопрягающим ионом. Основным сопрягающим ионом в клетке эукариот является Н+, и соответственно DmН+ является основной конвертируемой формой запасания энергии. Вторым по значимости сопрягающим ионом является Na+ (DmNa+). В то время как Сa2+, K+ и Cl- не используются для совершения какой-либо работы.
Биологическое окисление – это процесс дегидрирования субстрата с помощью промежуточных переносчиков водорода и его конечного акцептора. Если в роли конечного акцептора выступает кислород, процесс называется аэробным окислением или тканевым дыханием, если конечный акцептор представлен не кислородом – анаэробным окислением. Анаэробное окисление имеет ограниченное значение в организме человека. Основная функция биологического окисления – обеспечение клетки энергией в доступной форме.
Тканевое дыхание – процесс окисления водорода кислородом до воды ферментами цепи тканевого дыхания. Оно протекает по следующей схеме:
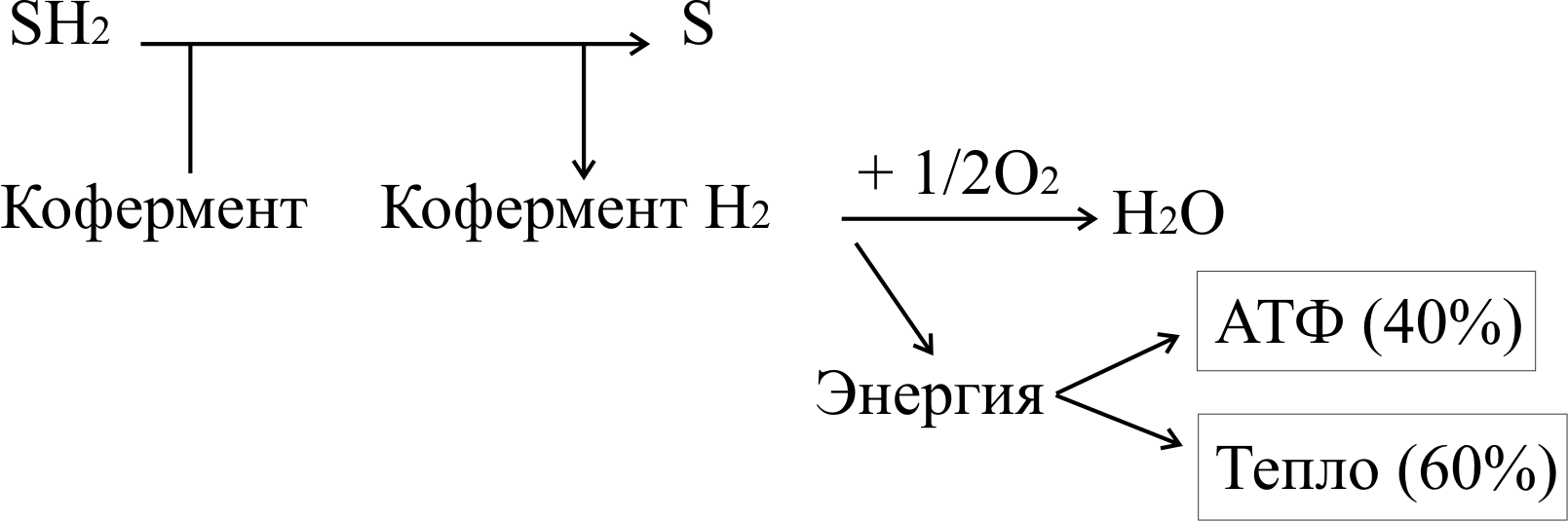
Вещество окисляется, если отдает электроны или одновременно электроны и протоны (атомы водорода), или присоединяет кислород. Способность молекулы отдавать электроны другой молекуле определяется окислительно-восстановительным потенциалом (редокс-потенциалом). Любое соединение может отдавать электроны только веществу с более высоким окислительно-восстановительным потенциалом. Окислитель и восстановитель всегда образуют сопряженную пару.
Выделяют 2 типа окисляемых субстратов:
Пиридинзависимые – спиртовые или альдегидные – изоцитрат, α-кетоглутарат, пируват, малат, глутамат, b-гидроксиацил-КоА, b-гидроксибутират, – в их дегидрировании участвуют НАД-зависимые дегидрогеназы.
Флавинзависимые – являются производными углеводородов – сукцинат, ацил-КоА, глицерол-3-фосфат, холин – при дегидрировании передают водород на ФАД-зависимые дегидрогеназы.
Цепь тканевого дыхания – последовательность переносчиков протонов водорода (Н+) и электронов от окисляемого субстрата на кислород, локализованных на внутренней мембране митохондрий.
| S1H2 НАД+ ФМН KoQ Fe3+ Fe3+ Fe3+ 1/2O2 S1 НАДH+H+ ФМНН2 KoQН2 Fe2+ Fe2+ Fe2+ H2O 2H+ АТФ АТФ АТФ |
| S2Н2 S2 ФАД ФАДН2 |
| ц и т о х р о м ы в с1с аа3 |
Рис. 10.1. Схема ЦТД
Компоненты ЦТД:
НАД-зависимые дегидрогеназы дегидрируют пиридинзависимые субстраты и акцептируют 2ē и один Н+.
ФАД (ФМН) - зависимые дегидрогеназы акцептируют 2 атома водорода (2Н+ и 2ē). ФМН – зависимая дегидрогеназа дегидрирует только НАДН, в то время как ФАД- дегидрогеназы окисляют флавинзависимые субстраты.
Жирорастворимый переносчик убихинон (кофермент Q, КоQ) – свободно перемещается по мембране митохондрий и акцептирует два атома водорода и превращается в КоQH2 (восстановленная форма – убихинол).
Система цитохромов – переносит только электроны. Цитохромы железосодержащие белки, простетическая группа которых по структуре напоминает гем. В отличие от гема атом железа в цитохроме может обратимо переходить из двух – в трехвалентное состояние (Fe3+ + ē « Fe2+). Это и обеспечивает участие цитохрома в транспорте электронов. Цитохромы действуют в порядке возрастания их редокс-потенциала и в дыхательной цепи располагаются следующим образом: b - с1 – с – а - а3. Два последних работают в ассоциации как один фермент цитохромоксидаза аа3. Цитохромоксидаза состоит из 6 субъединиц (2 - цитохрома а и 4 - цитохрома а3). В цитохроме а3 кроме железа имеются атомы меди и он передает электроны непосредственно на кислород. Атом кислорода при этом заряжается отрицательно и приобретает способность взаимодействовать с протонами с образованием метаболической воды.
Железосерные белки (FeS) – содержат негемовое железо и участвуют в окислительно-восстановительных процессах, протекающих по одноэлектронному механизму и ассоциированы с флавопротеинами и цитохромом b.
Дата добавления: 2015-10-05; просмотров: 1210;