Эмбриональная регуляция
Выше было показано, что эксперименты по выявлению сроков детерминации в раннем развитии разных видов животных привели к открытию явления эмбриональной регуляции, т.е. процесса восстановления нормального развития целого зародыша или зачатка После его естественного или искусственного нарушения. Опыты, многочисленные и очень разнообразные, дали интереснейшие результаты.
Для изучения регуляционных возможностей зародышей использовали следующие методические приемы: 1) удаление части материала зародыша; 2) добавление избыточного материала; 3) перемешивание материала, а также изменение пространственного взаиморасположения путем центрифугирования или сдавления. Эти манипуляции проводили на стадиях яйца, зиготы, дробления, гаструляции и органогенеза.
До начала дробления от яйцеклеток отбирали часть цитоплазмы путем отсасывания пипеткой или отрыва в результате сильного центрифугирования. Обнаружено, что за очень редкими исключениями изъятие даже 1/3 цитоплазмы яйца не препятствует дальнейшему нормальному развитию зародыша, хотя и меньшего размера. Такие результаты были получены на яйцеклетках кишечнополостных, многощетинковых червей, моллюсков, асцидий, иглокожих. Слияние двух или нескольких яйцеклеток тоже заканчивалось нормальным развитием.
Если яйца, выпущенные из гонад морского ежа, поместить в тесный сосуд, то отдельные яйца сливаются и образуются гигантские яйца. После искусственного оплодотворения такого яйца из него развивается совершенно нормальная личинка, но вдвое больших размеров. Количество клеток в ней такое же, как и у обычной личинки, но каждая клетка вдвое крупнее. Таким образом, две овоплазмы, сливаясь вместе, образуют одно органическое целое.
Опыты по центрифугированию яиц тоже дали интересные результаты. Показано, что сильное центрифугирование яиц моллюсков, червей и морского ежа не изменяет места выделения полярных телец, направления первой борозды дробления и характера развития. Получается совершенно нормальная личинка со случайным распределением пигмента и других веществ. Яйцо тритона после 20-минутного центрифугирования тоже дробится, несмотря на полное нарушение расположения в нем желтка и других компонентов. Существует предположение, что определенное размещение разных сортов цитоплазмы внутри яйца и зиготы не есть причина всего дальнейшего развития, а наоборот, само пространственное расположение цитоплазмы является следствием других, более глубоких причин, которые нужно искать в энергетике и физиологии яйца.
Все приведенные опыты показывают, что овоплазматическая сегрегация не является абсолютной причиной детерминации и при ее нарушении возможна эмбриональная регуляция.
Множество опытов проведено в стадии дробления. Почти у всех видов животных разделение бластомеров на первых этапах дробления приводит к развитию из них целых, нормальных зародышей, т.е. монозиготных близнецов. У морского ежа каждый из четырех бластомеров может образовать нормальную личинку. На стадии восьми бластомеров полноценная регуляция осуществляется в бластомерах вегетативного полюса, а в бластомерах анимального полюса она возможна при помещении их в слабый раствор солей лития.
Регуляция во время дробления широко распространена у позвоночных: миног, хрящевых и костных рыб, земноводных и высших позвоночных. Об этом свидетельствуют опыты по изоляции бластомеров и частое образование монозиготных близнецов и двойниковых уродств. Возможно развитие нормальных крольчат из одного удаленного бластомера двух-, четырех- и восьмиклеточной стадии зародыша в 30, 19 и 11% случаев соответственно. У четырехклеточного зародыша мыши при разделении бластомеров возможно развитие трех бластоцист и одного трофобластического пузырька. Даже у нематод, являющихся образцом мозаичного дробления, обнаружено, что изолированный бластомер с половым зачатком может образовать цельный зародыш. Из этих опытов следует, что потенции отдельного бластомера к морфогенезу шире, чем реализующиеся из него в ходе нормального развития зачатки. Потенции —это максимальные возможности элементов зародыша, т.е. направления их развития, которые могли бы осуществиться. В норме реализуется лишь одно из них, а остальные могут быть выявлены в эксперименте. Широкие потенции называют еще тотипотентностью.
Широкие потенции бластомеров подтверждаются многочисленными опытами по их пересаже и перемешиванию. Бластомеры морского ежа разделяли и затем перемешивали в беспорядке. Получились нормальные личинки, но другим путем: кишечник, например, образовывался не инвагинацией, а шизоцельно (путем расхождения) из плотной клеточной массы; скелет возникал раньше, чем покровы. Это пример эквифинальности, которую можно рассматривать как одно из важнейших свойств регуляционных процессов.
У зародышей млекопитающих бластомеры при пересадке ведут себя соответственно месту пересадки. Это подтверждено переносом клеток из трофобласта в зародышевый узелок, и наоборот. В этих случаях проявляются зависимость судьбы бластомера от взаимодействия с другими бластомерами и более широкие его потенции. В таком же ключе можно интерпретировать следующие опыты: если бластоцисту поместить в полость тела, то из нее развивается солидная опухоль (эмбриокарцинома). Клетки этой опухоли, будучи введены в бластоцисту, приводят к нормальному развитию зародыша.
Большой интерес представляют опыты по объединению зародышей. Искусственное слияние двух зародышей морского ежа на стадии бластулы может завершиться формированием совершенно нормальной структуры личинки. Такая личинка превосходит по размерам нормальных зародышей и имеет вдвое больше клеток. Это доказывает, что при совершенно тождественной архитектонике тела оно может быть построено из разного количества клеток. Следовательно, детерминация носит не целлюлярный характер, а является производной от организма как целого.
Слияние двубластомерных зародышей возможно получить у тритона.
В отношении млекопитающих было высказано предположение, что в их зародышах предетерминированные локализованные участки цитоплазмы не играют никакой роли. Зависимость судьбы клетки от ее положения была продемонстрирована следующим образом. Объединяли клетки двух мышиных зародышей, находящихся на стадии морулы и предварительно диссоциированных. Зародыши отличались по генам окраски шерсти. Образовавшуюся в результате гибридную бластоцисту имплантировали в приемную мать. Развился нормальный живой мышонок-химера, в окраске которого проявилось действие обоих генов обоих родителей (рис. 8.16).
Как было отмечено выше, в яйцах всех животных цитоплазма неоднородна. Она постепенно изменяет свои свойства вдоль главной оси яйца. Фактически у всех животных до дробления детерминируются полярность, симметрия, дорсовентральная ось и тип дробления. Однако нет основания понимать структуру цитоплазмы яйца в преформационном духе, т.е. думать, что в ней уже имеются в готовом виде органообразующие зачатки и что она столь же сложно сконструирована, как и взрослый организм. Об этом, в частности, свидетельствуют опыты по сдавливанию зародышей в стадии дробления. Если яйцо лягушки сдавить двумя пластинками в анимально-вегетативном направлении, то после двух меридиональных дроблений, происходящих, как и при нормальных условиях, следует не горизонтальное, а опять меридиональное, приводящее к расположению восьми бластомеров в один слой в виде пластинки. То же наблюдается и на стадии 16 и 32 бластомеров. После удаления давящей силы из таких яиц могут развиться нормальные зародыши. Судьба бластомеров в этом случае, конечно, иная, чем при нормальном развитии.
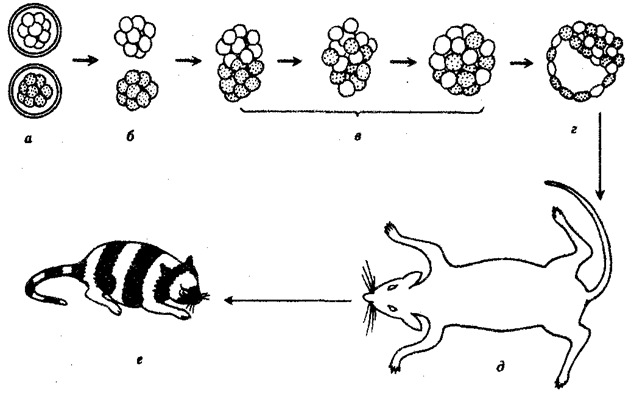
Рис. 8.16. Химерный мышонок, полученный в результате объединения
на стадии морулы двух зародышей с разными генотипами:
а—дробящиеся яйца, б—морулы, освобожденные от яйцевых оболочек путем обработки протеолитическими ферментами, в—объединяющиеся морулы, инкубированные in vitro при 37°С, г—объединенный зародыш на стадии бластоцисты, д—мать-реципиент с имплантированным химерным зародышем, е—химерный мышонок
Наличие периода развития, когда потенции элементов зародыша шире, чем реализующиеся при обычном развитии, т.е. когда детерминация не носит окончательного, необратимого характера, лежит в основе эмбриональных регуляций. Эмбриональные регуляции были открыты немецким эмбриологом Г.Дришем (1908). Таким образом, детерминация и эмбриональная регуляция являются противоположными свойствами и теснейшим образом взаимосвязаны в системе целостного развивающегося зародыша.
Детерминация, как правило, идет от целого к частям: сначала детерминируется целый зачаток зародыша, но судьба отдельных его элементов (клеток) еще не определена, затем постепенно или скачкообразно детерминируются отдельные элементы. Обычно детерминация сменяется дифференциацией и специализацией частей организма, усиливается взаимная зависимость их друг от друга, возрастают интегрированность, целостность организма. Исключение составляют виды, у которых взрослые формы устроены проще личинок. Способность к эмбриональной регуляции в ходе онтогенеза соответственно падает, но не абсолютно, так как и у взрослого организма существует, например, способность к регенерации, т.е. к восстановлению утраченных или поврежденных частей.
Важно, что не только в эксперименте, где искусственно нарушается нормальный ход развития, но и в естественных условиях ход развития клеток регулируется согласно их положению в целом.
Вероятно гибкий регуляторный характер развития имеет существенные адаптивные преимущества: если какое-либо одно звено будет подавлено или полностью выпадает, сохраняются шансы на достижение нормального конечного результата. С другой стороны, в экстремальных условиях резерв изменчивости может стать источником эволюционных преобразований. Эти предположения вполне согласуются с фактом регуляционного типа яиц у позвоночных и с тем, что вообще способность к эмбриональной регуляции у них выражена сильнее, чем у многих беспозвоночных.
Дата добавления: 2015-09-07; просмотров: 816;