Зрительная предметная агнозия и нарушения переработки информации в мозге
Роль переработки информации об общих свойствах объекта(см. обзор в Ulman, 1996; Тагг, Bulhoff, 1999). Прямой подход к распознанию объектов основан на допущении, что образ воспринимаемого объекта может быть распознан при его сравнении с образами объектов, хранящихся в памяти. Однако число образов даже единственного объекта, отраженного с разных точек зрения и при разных углах падения света, было бы слишком велико и требовало бы огромного объема памяти. Кроме того, образ, который требуется распознать, может несколько отличаться от всех образов, виденных в прошлом.
Было бы разумно предположить, что система распознания объектов была бы намного более эффективной (в смысле экономии объема памяти и надежности распознания конкретного объекта), если бы она опиралась на переработку общих свойств объекта, число которых ограничено и которые намного легче сравнивать с небольшим количеством образов памяти (Ullman, 1996). Чтобы распознать стол, необязательно хранить в памяти большое количество образцов форм, текстур, месторасположений и расцветок разнообразных столов в различных ракурсах. Общие свойства стола любой формы, местоположения, расцветки, текстуры или размера могут храниться в памяти и использоваться для сравнения с соответствующими свойствами распознаваемого объекта.
Стадии переработки зрительной информации.Исследования приматов показывают, что переработка информации об общих свойствах объектов происходит в многочисленных специализированных и взаимосвязанных областях зрительной системы и включает несколько последовательных стадий. Предполагается, что эти стадии таковы:
1) предварительная переработка элементарных характеристик зрительной информации, таких как освещенность, цвет, движение;
2) промежуточная переработка — группировка элементарных характеристик с похожими свойствами в объекты и их общие очертания;
3) окончательная переработка, приводящая к восприятию объектов и сцен на основании результатов промежуточной переработки (Van Essen et al., 1990).
Зрение может пострадать при нарушении любой из трех стадий, но развитие зрительной агнозии объектов может быть связано главным образом со второй и особенно с третьей стадией переработки зрительной информации. Нарушения на стадии окончательной переработки зрительной информации — восприятия объектов — описаны в предыдущих разделах. Нарушения предварительной и промежуточной переработки и их связь со зрительной предметной агнозией рассматриваются в следующих разделах.
Нарушения низшего уровня переработки зрительной информации и зрительная предметная агнозия.Частичная потеря элементарной зрительной чувствительности. Некоторые авторы подчеркивают исключительную роль нарушений элементарной зрительной чувствительности при развитии зрительной агнозии. Бэй предполагает, что легкие нарушения зрительной чувствительности могут объяснять расстройства идентификации объектов (Bay, 1950; 1953). Он утверждает, что динамические процессы могут выявить такие элементарные нарушения, и обнаружил у пациентов со зрительной агнозией расстройства локальной адаптации в нормальных областях полей зрения. По его мнению, стимул," поступивший на эти поля, у таких пациентов угасает намного быстрее, чем у здоровых испытуемых. Бэй подчеркнул роль слабоумия и сниженного уровня сознания как дополнительного фактора развития зрительной агнозии у пациентов с нарушениями элементарной зрительной чувствительности.
Эттлингер указал на то, что элементарные нарушения зрения можно обнаружить у многих пациентов, не страдающих зрительной агнозией (Ettlinger, 1956). Он использовал обширную батарею тестов элементарных зрительных процессов, в том числе различения яркости, слияния мельканий, резкости восприятия мелких объектов, тахистоскопической резкости, локальной адаптации, и не обнаружил значимой корреляции между нарушениями элементарных сенсорных процессов и зрительной агнозией у 30 пациентов с поражениями мозга. Аналогичные результаты были получены Кантимулиной, которая обнаружила расстройства различения яркости и локальной адаптации у пациентов с затылочными поражениями при отсутствии зрительной агнозии (Кантимулина, 1961). Конечно, значительное снижение остроты зрения ухудшает способность выделять элементарные зрительные свойства, но такие нарушения можно преодолеть на последующих стадиях переработки зрительной информации.
Нарушения на промежуточном уровне переработки зрительной информациии зрительная агнозия объектов.Нарушения на уровне анализа простых свойств. Мозговые корреляты процесса анализа простых свойств. Со времен классических работ Хьюбела и Визеля известно, что ранние стадии переработки формы, в том числе ориентации линий, их пересечений и углов, у приматов проходят в области V1 (поле 17 Бродмана) (Hubel, Wiesel, 1959; 1968; 1977). Также обнаружено, что ранние стадии зрительной переработки элементов, которые можно использовать при анализе формы, локализуются в области V2. Эти области подготавливают информацию для переработки на высшем уровне и, таким образом, отвечают за ее промежуточную переработку.
В области V1, которая находится в стриарной коре, соответствующей полю 17 Бродмана, расположена серия капель и межкапельных участков. Нейроны области VI чувствительны к свету в пределах рецептивного поля, которое получает стимулы от небольшого участка сетчатки одного или обоих глаз через латеральные коленчатые ядра. В области V1 имеются ориентационные колонки и колонки глазодоминантности. Это было впервые показано в классической работе Хьюбела и Визеля (Hubel, Wiesel, 1959; 1968; 1977). В каждой ориентационной колонке представлена определенная ориентация линий. Среди других функциональных свойств колонок области V1 можно назвать ретинотопию, глазодоминантность
и цвет. Клетки рецептивного поля V1 реагируют на ориентацию линий, величину угла, контрастные границы контура, ориентацию контура, пересечение линий, движение и бинокулярную диспаратность. Рецептивные поля со слепыми окончаниями равно предпочитают линии и пересечения, но лучше всего реагируют на короткие линии и короткие фрагменты углов (см. обзор в von der Heudt, 1995). У обезьян клетки со слепыми окончаниями можно обнаружить в областях V1 и V2. Некоторые клетки в области капель получают закодированный в цвете сигнал от клеток латеральных коленчатых ядер.
Область V2 расположена в престриарной коре, занимает часть поля 18 Бродмана и содержит различные паттерны «толстых», «тонких» и «бледных» полосок («thick dense», «thin dense», «pale bands»). Области V1 и V2 во многом взаимосвязаны и являются дивергентными и конвергентными (см. обзор в Ts'o & Roe, 1995). Стимулы из V2 передаются в область V4, а затем в кору средневисочной области. Рецептивные поля в области V2 больше, чем в V1, и ретинотопия в V2 менее точная, чем в V1. Нейроны области V2 реагируют на более интегрированные свойства, например, клетки диспаратности реагируют на оптимальную стереоскопическую диспаратность сетчатки двух глаз, но не откликаются на монокулярную стимуляцию. Как и нейроны области VI, нейроны V2 активируются ориентацией, но, кроме того, они могут реагировать на контрастные границы как реальных, так и воображаемых, «субъективных» контуров (von der Heydt et al., 1984; von der Heydt, Peterhans, 1989). Эти нейроны расположены в районе тонких бледных полос области V2 (см. Grusser, Landis, 1991; von der Heydt, 1995). Нейроны «тонких полос» реагируют на хроматические стимулы. Область V2 (часть поля 18 Бродмана) содержит нейроны, которые характеризуются сильной настройкой на ориентацию и чередование светлых и темных полос, что указывает на их способность к избирательной пространственно-частотной настройке, являющейся первым этапом распознания текстуры и следующим за областью VI этапом определения контуров (см. обзор в Grusser, Landis, 1991).
Нарушения на уровне анализа простых свойств или нарушения различения простых свойств.Сохранность дифференциальных порогов при неограниченном времени предъявления и коротком межстимульном интервале. Рассмотренные данные указывают на возможность того, что распознание ориентации линий, величины углов, текстур может проходить на ранних стадиях восприятия объекта в мозге. Обнаружено, что можно выявить нарушения в способности воспринимать наклон двух линий как одинаковый или разный (Warrington, Rabin, 1970). Были разработаны специальные тесты для диагностики способности пациентов подбирать наклон линии по имеющемуся образцу, где они расположены в виде «солнечных лучей» и образуют с горизонталью углы от 0° до 180° с шагом в 16°. Длина линии «луча» и стимула одинакова. В усложненном варианте теста сти-мульная линия была значительно короче «луча» (Benton et al, 1978).
Данные нарушения могут играть некоторую роль в развитии зрительной агнозии объектов. Предположив это, мы ожидали обнаружить у пациентов со зрительной агнозией объектов и поражениями затылочной доли различную статистическую частоту нарушений распознания ориентации линий, различения величины углов и текстур. Однако при времени предъявления стимула 2000 мс и межсти-
мульном интервале 2 с не было обнаружено значительных нарушений. Трудности начали возникать, когда время предъявления сокращалось до 200 мс либо меж-стимульный интервал увеличивался до 5 или 10 с.
Мы обследовали 32 пациента после инсульта (Тонконогий, 1973; Меерсон, 1986) и 42 пациента, перенесших удаление передненижней височной доли в ходе операции по лечению височной эпилепсии (Меерсон, 1977; Меерсон, 1986). В группе пациентов после инсульта зрительная агнозия объектов и возможные поражения затылочно-височной области были обнаружены у 8 человек, в том числе билатеральные поражения у 4 человек и левотеменные поражения у 4 человек. В контрольную группу входили следующие пациенты:
1) с конструктивной апраксией и преимущественно нижнетеменными поражениями — три случая с левосторонними и один случай с правосторонними поражениями;
2) с афазией Вернике и верхнезадними височными поражениями — 12 случаев, все левосторонние;
3) с афазией Брока, левосторонней или правосторонней гемиплегией и фронтальными премоторными поражениями — 8 случаев, из них 6 пациентов с левосторонними и 2 пациента с правосторонними поражениями.
В число клинических критериев локализации поражений также входило наличие гемианопсий и типы гемианопсии. Пациенты тестировались в период от 5-6 недель до нескольких месяцев после инсульта.
В группу пациентов с удалением передненижней височной доли входили 22 человека, перенесшие операцию на правом полушарии, и 20 человек — на левом полушарии. У них были удалены такие части передненижней височной доли, как парагиппокампальная и веретенообразная извилина и медиобазальные структуры, прежде всего гиппокамп. В этой группе пациентов зрительная агнозия" объектов не обнаружена. Пациенты тестировались через 4-6 недель после операции (Меерсон, 1986, стр. 124-125). Различение углов и текстур исследовалось по тому же принципу, что и у пациентов с иной локализацией поражений.
Стимульный материал представлял собой линии, отклоняющиеся от горизонтали на 30-75°, и углы такой же величины (рис. 2.1.5). В исследовании пациентов, перенесших инсульт и операцию, величина углов и отклонений линий составляла 5-45° и 1-45° соответственно.
В другой набор стимулов входили текстуры с разными статистическими характеристиками. Текстуры состояли из маленьких черных квадратиков, выстроенных в линии, на фоне большого белого квадрата (Julezs, 1971). Они различались по частоте маленьких черных квадратиков, изображенных в случайном порядке в соответствии со статистикой первого порядка в диапазоне отр = 0,1 дор = 0,9, с разностями от р = 0,05 до р = 0,4 (см. рис. 2.1.5).
Стимулы предъявлялись на экране тахистоскопа со временем экспозиции 2 с. Интервал между двумя сравниваемыми стимулами составлял 0 с, то есть два стимула предъявлялись одновременно. Увидев пару стимулов, испытуемый должен был сказать, являются ли, например, два угла одинаковой или разной величины одинаковыми или разными. После завершения теста подсчитывались средние
дифференциальные пороги. Мы ожидали, что возможные нарушения в областях VI и V2, исследованные Хьюбелом и Визелем, проявятся в увеличении дифференциальных порогов различения углов и текстур.

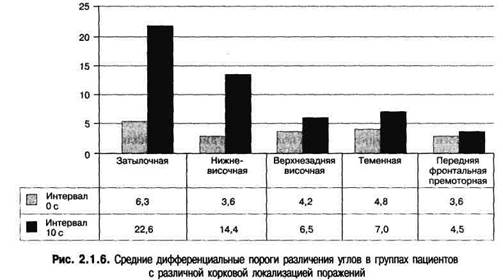
Рис.2.1.5. Примеры углов и текстур, используемых для дифференциации двух стимулов. Вероятность черных квадратов для верхней текстуры - р = 0,2, для нижней - р = 0,45
Результаты различения углов указывают на отсутствие значимых различий дифференциальных порогов в двух группах пациентов — со зрительной агнозией объектов и затылочными или затылочно-височными поражениями и с различной локализацией поражений вне затылочной области (Меерсон, 1986). При времени экспозиции 2000 мс и одновременном предъявлении двух стимулов для различения средние дифференциальные пороги различения углов и текстур были одинаковыми во всех группах пациентов (рис. 2.1.6). Пороги различения углов были чуть выше в группе с затылочными поражениями и составляли 6,3 по сравнению с 4,8 у пациентов с теменными поражениями, 4,2 у пациентов с верхнезадними височными поражениями и 3,6 у пациентов с премоторными фронтальными поражениями. Аналогичные результаты — 3,6 — были получены в группе с нижневисочными поражениями.
Дизайн исследования: два угла, изображенных внизу рисунка, предъявлялись на экране тахистоскопа с интервалом 0 с (одновременно) или 10 с. Время предъявления — 2 с. Испытуемый должен сказать, одинаковые эти стимулы или разные.
Как и в эксперименте с различением углов, средние дифференциальные пороги различения текстур при одновременном предъявлении не различались значимым образом у пациентов с затылочными, теменными, верхневисочными, задне-височными и премоторными фронтальными поражениями и составляли 0,08; 0,075; 0,071; 0,085 и 0,065 соответственно.
Эти результаты указывают на возможную сохранность промежуточных стадий переработки простых свойств, таких как различение углов и текстур, у пациентов с различными типами локальной патологии мозга. В том числе у пациентов с наличием или отсутствием затылочных поражений, при условии предъявления сигналов в конвенциональном виде с оптимальным временем экспозиции и меж-стимульными интервалами в 0, 5 и 10 с. Некоторое улучшение различения углов, отмеченное в группе с затылочными поражениями по сравнению с другими группами пациентов, возможно, связано с довольно незначительными нарушениями в оценке величины углов на низшем уровне переработки зрительной информации. Различия между группами пациентов с различной локализацией поражений стали более значимыми при увеличении межстимульных интервалов или уменьшении времени экспозиции.
Увеличение межстимульных интервалов. Расстройства зрительной рабочей памяти. Во второй серии экспериментов по различению углов время экспозиции стимулов осталось 2000 мс, но интервалы между двумя стимулами варьировались: 0 с, 5 с и 10 с. Результаты представлены в табл. 2.1.2. При одновременном предъявлении стимулов средние дифференциальные пороги для углов были немного выше в группе с затылочными поражениями, чем в других группах пациентов, но эти различия значительно усилились при увеличении межстимульных интервалов. Например, средние дифференциальные пороги различения углов в группе с теменными поражениями возросли от 4,6 до 8,9 при интервалах 0 и 10 с соответственно, а в группе с затылочными поражениями произошло возрастание от 6,3 до 22,6. Значительное увеличение средних дифференциальных порогов различения углов также обнаружилось в группе пациентов с передневисочными поражениями: от 3,6 при межстимульном интервале 0 с до 14,4 при интервале 10 с (см. табл. 2.1.2).
Таблица 2.1.2 Изменения дифференциальных порогов распознавания углов и текстур при увеличении межстимульного интервала от 0 с (одновременное предъявление) до 5 с и 10 с
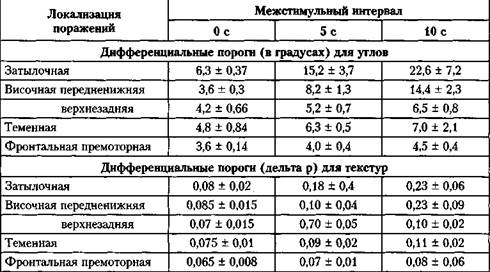
Аналогичные результаты были получены по дифференциальным порогам различения текстур (см. табл. 2.1.2). С увеличением межстимульного интервала от 0 до 10 с средние дифференциальные пороги в группе с затылочными поражениями возросли почти в три раза — с 0,08 до 0,23, в группе с передненижними височными поражениями — с 0,085 до 0,23, а в остальных группах возросли лишь незначительно.
Кратковременная экспозиция. Процедура тестирования не отличается от той, которая применялась при увеличении межстимульных интервалов, только эти интервалы фиксировались на 2 с, а время экспозиции сократилось с 2000 до 500 и 200 мс. В группе с затылочными поражениями это привело к заметному возрастанию дифференциальных порогов от 6,3 ± 0,37 до 9,6 ± 0,85 при времени экспозиции 500 мс и до 13,3 ± 2,6 при времени экспозиции 200 мс. В остальных группах пациентов значимого возрастания дифференциальных порогов не произошло. Например, в группе с верхнезадними височными поражениями дифференциальные пороги различения углов составляли 5,3 ± 0,3 при 2000 мс, 5,5 ± 0,66 при 500 мс и 6,1 ± 0,18 при 200 мс. Группа с передненижними поражениями не включалась в тестирование.
Результаты тестов можно объяснить возможными нарушениями некоторых операций, проходящих в колонках нейронов, расположенных в областях VI и V2. Возможно, операции, за которые отвечают эти нейроны, частично или полностью отвечают за сохранение информации об ориентации линий, величине углов и свойствах текстур в зрительной рабочей памяти для дальнейшей переработки. Поэтому операции могут нарушаться, когда рабочая память нагружается за счет увеличения межстимульного интервала или уменьшения времени экспозиции. Это объ-
яснение может быть основано на роли высоких и низких частотных характеристик пространства при переработке информации. Возможно, информация низкой пространственной частоты лучше сохраняется и менее подвержена нарушениям, а информация высокой пространственной частоты более уязвима и менее защищена. Поэтому дифференциальные пороги увеличиваются в трудных неконвенциональных условиях при кратковременном предъявлении или увеличении межстимульного интервала с сохранением низкочастотных градаций сигнала и нарушением высокочастотных различий градации.
Операции, участвующие в подготовке зрительной информации для дальнейшей переработки, могут быть направлены на сохранение всего объема информации путем ее объединения или группировки в большие группы. Возможный механизм такого уменьшения объема информации может быть связан с процессом разбора стимула на составляющие для его дальнейшей переработки с использованием фильтров высоких и низких пространственных частот. В рабочей памяти пациентов с корковой патологией разбор стимула на небольшие составляющие с помощью высокочастотных фильтров может нарушаться, хотя разбор его на большие части с помощью низкочастотных фильтров остается сохранным. Например, углы после первоначального запоминания могут быть объединены в большие группы с разницей по величине в 15°. Это позволяет разбить углы от 5° до 45° на три большие группы, переработать которые легче, чем 9 маленьких групп с разницей по величине в 5° (см. табл. 2.1.2). При разбиении на большие группы в рабочей зрительной памяти требуется меньше места для хранения информации и облегчается ее защита от воздействий, что очень важно при переработке хорошо знакомой зрительной информации конвенционального типа. В то же время, разбиение на многочисленные маленькие группы занимает больше времени и больше места в хранилище рабочей памяти. Но маленькие группы могут быть полезны в случае переработки неконвенциональной, новой информации.
Увеличение дифференциальных порогов различения углов и текстур у пациентов с затылочными поражениями ограничивается зрительной модальностью. При различении звуковых тонов увеличение межстимульного интервала не привело к значимому возрастанию дифференциальных порогов у пациентов с затылочными поражениями, хотя в группе пациентов с передневисочными поражениями было зафиксировано значительное увеличение порогов в зрительной и слуховой модальности (см. раздел 2.5 «Слуховая агнозия»). Эти результаты указывают на возможные различия, лежащие в основе процессов группировки информации в рабочей памяти, происходящих в затылочной и передненижней височной областях. Возможно, зрительная информация о простых свойствах объекта анализируется в зрительной рабочей памяти в затылочной области для использования на последующих стадиях переработки зрительной информации. В то же время, анализ информации в зрительной рабочей памяти в затылочной области и в слуховой рабочей памяти в верхнезадних височных областях используется для перевода информации из одномодальных слуховых или зрительных кодов в многомодальный код в передненижних областях. Перевод информации, объединенной в большие группы, может сохраняться, но перевод информации, разбитой на многочисленные маленькие группы, нарушается, что приводит к увеличению дифференциальных порогов при поражениях передненижних височных областей.
Нарушения процессов группировки в рабочей памяти и сохранность распознания объектов после удаления передненижней височной доли указывают на возможность того, что нарушения этих процессов не играют важной роли при развитии зрительной агнозии объектов или могут служить дополнительным фактором, осложняющим уже существующие нарушения высшего уровня переработки зрительной информации. Это выступает особенно отчетливо в ходе запоминания или распознания объектов в неконвенциональном виде или других типов переработки неконвенциональной зрительной информации.
Механизмы, лежащие в основе анализа стимулов при кратковременном предъявлении, также могут быть связаны со зрительной рабочей памятью в том случае, когда предъявление удлиняется, чтобы обеспечить время для анализа стимулов, необходимого для дальнейшей переработки. Увеличение времени предъявления может способствовать анализу ограниченного числа больших групп зрительной информации, но может быть несущественным для переработки многочисленных мелких групп, что приводит к увеличению дифференциальных порогов.
Нарушения кодирования простых свойств объектов при кратковременном предъявлении могут играть некоторую роль в развитии зрительной агнозии объектов. Возможно, распознание объектов с помощью более точного описания их простых свойств может нарушаться при неспособности пациентов завершить эту операцию за короткий промежуток времени. Но это может главным образом относиться к неконвенциональному типу зрительной агнозии объектов, поскольку конвенциональная агнозия объектов обычно проявляется при неограниченном времени предъявления.
В то же время, можно наблюдать больше нарушений в различении ориентации линий, величины углов и текстур при неограниченном времени предъявления, когда пациентам со зрительной агнозией объектов приходится различать эти относительно простые свойства как часть более сложных свойств.
Нарушения высших уровней переработки зрительной информации и зрительная предметная агнозия.Нарушения на уровне анализа сложных свойств. Мозговые корреляты процесса анализа сложных свойств. Область V4 включает в себя престриарную зону зрительной коры, в том числе, участки веретенообразной и язычной борозды (McKeefy, Zeki, 1997) и может быть связана с высшим уровнем процессов распознания объекта. Нарушения областей V2 или V4 у животных также влияют на способность различать объекты в плотном потоке стимулов, «соревнующихся» друг с другом в интенсивности (Schiller, 1993).
Из области V4 стимулы поступают в нижневисочную область (НВ), которая включает в себя заднюю нижневисочную зону (ЗНВ) и переднюю нижневисочную зону (ПНВ). Билатеральное удаление НВ у обезьян приводит к постоянным нарушениям зрительного различения и распознания (Gross, 1992; Ungerleider, Mish-kin, 1982). Психологические исследования показывают (см. обзор Young, 1995), что клетки ПНВ сильнее реагируют на сложные стимулы, чем на простые линии или точки (Tanaka et al, 1990; Fujita et al., 1992). Некоторые клетки отвечают на специфические стимулы, такие как руки (Gross et al., 1992) или лица (Bruce etal., 1981; Desimone et al., 1984; Young, Yamane, 1992), но нечувствительны к расположению, размеру, цвету или пространственной частоте стимула. Одна группа кле-
ток, избирательно реагирующих на лица, отвечает на целостные образы лиц, другая — на части лица.
Некоторые клетки в НВ области предпочитают конкретное расположение объекта. Определенная ориентация объекта вызывает более сильный ответ. Другие клетки широко настроены на многочисленные ракурсы объекта, возможно, получая конвергирующую информацию со многих клеток, настроенных на конкретное расположение (Logothetis, 1995). Это может быть связано с применением позиционно-комбинирующего подхода (view-combination approach) (Ulman, 1999).
Показано, что простые стимулы в виде очень похожих элементов разнообразных объектов могут возбуждать определенные области, прилегающие к соседним нейронам (Fujita et al., 1992). Нейроны, организованные в виде колонок или модулей в вертикальном разрезе, отвечают на один и тот же стимул так же, как самый первый нейрон. Эти данные позволяют предположить, что нейроны работают в качестве детекторов фрагментов паттернов, или «алфавита» паттернов, благодаря чему играют основную роль в процессе распознания (Stryker, 1992). Однако нейрон не отвечает на разнообразные стимулы, включающие в себя предпочитаемый им фрагмент паттернов. Таким образом, наличие одной из «букв» алфавита в объекте или фигуре не возбуждает соответствующий нейрон. Это позволяет заключить, что, по крайней мере, предложенный исследователями алфавит основных свойств не соответствует алфавиту, хранящемуся в мозге обезьян.
Подробный обзор данных, полученных при простой записи активности нейронов, представлен в Grusser, Landis (1991), Ts'o, Roe (1995), Parker et al. (1995). Эти данные представляют собой важный шаг в понимании механизмов анализа формы и распознавания объектов в зрительной системе. Однако остается неясным, каким образом процесс переработки формы переходит от распознания простых свойств в областях V1 и V2 к распознанию сложных свойств и формы объектов в НВ области. Этот процесс включает в себя множество сложных операций, связанных с переработкой зрительной информации конвенционального и неконвенционального типа, которые отражены в современных теориях, основанных на психологических и нейропсихологических исследованиях и моделировании процесса зрительного распознания объектов. Конечно, требуются дальнейшие исследования по записи активности нейронов. Также было бы очень интересно рассмотреть очевидные противоречия между реакцией НВ клеток на визуально предъявленные объекты и анатомическими данными о том, что при зрительной агнозии объектов поражения преимущественно локализуются в затылочной и задневисочно-затылочной областях.
Нарушения формирования сложных свойств из простых свойств.Различение сложных свойств. Диагностировать способность различения сложных фигур, составленных из сочетания простых свойств, по одному из этих простых свойств можно с помощью набора простых свойств или отличающихся друг от друга сложных фигур. Один из таких тестов был разработан Эфроном (Efron, 1968). Автор исследовал различение формы, предъявляя испытуемому для сравнения пары прямоугольников. Все фигуры уравнены по яркости и площади (рис. 2.1.7).
Основная фигура (справа) сравнивается с фигурами слева. Фигуры в паре одинаковы, если длины их вертикальных и горизонтальных сторон равны. У одной

из фигур в разных парах форма оставалась постоянной, а у второй фигуры соотношение длин горизонтальной и вертикальной сторон постепенно увеличивалось. Испытуемый должен был сравнить это соотношение у прямоугольников, уравненных по простым свойствам, таким как яркость и площадь. От испытуемого требовалось сказать, одинаковые фигуры или разные.
Тест не представлял никаких трудностей для здоровых испытуемых. С помощью теста Эфрона Уоррингтон обследовал двух пациентов, Д. А. Ф. и Р. Б. С, со зрительной агнозией объектов и лиц (Warrrington, 1986). Обнаружено, что результаты тестов испытуемых зависели от степени сходства двух прямоугольников. Результаты значительно ухудшались, когда соотношение горизонтальной и вертикальной сторон становилось очень маленьким.
Со временем тест Эфрона стал стандартным
методом исследования различения формы у пациентов со зрительной агнозией объектов. Однако на самом деле он диагностирует различение сложных свойств прямоугольников, различающихся по одному из простых свойств. Было бы полезно сравнить результаты теста Эфрона с тестированием различения такого простого свойства, как соотношение длин горизонтальных и вертикальных линий, равных или различных, взятого отдельно от других свойств прямоугольников. Это поможет выявить нарушения процесса формирования сложных свойств, связанные с трудностями распознания простых свойств объекта. Такой тест был разработан одним из авторов (Тонконогий, 1973).
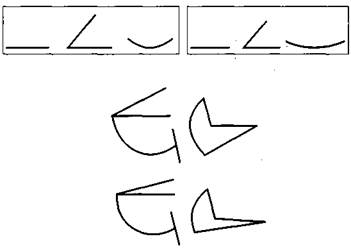
Рис. 2.1.8 А. Комплексы, состоящие из трех простых свойств - ориентация линии, величина угла
Дата добавления: 2015-09-18; просмотров: 1440;