A-кетоглутаратдегидрогеназным комплексом.
Регуляцию скорости ОПК осуществляя сколько механизмов:
1. аллостерическая регуляция - каждый регуляторный фермент имеет аллостерические эффекторы, концентрация кои изменяется в зависимости от состояния клетки;
2.увеличение активности фермента при высоких концентрации субстрата (например, пируват наиболее эффективный активатор пируватгидрогеназного комплекса);
3.ингибирование фермента продуктами peaкции: пируватдегидрогеназный комплекс инибируется ацетил-КоА и НАДН, цитратсинтаза - цитратом;
4. фосфорилирование и дефосфорилированиеПДК (рис.9).
Наиболее сложна регуляция ПДК. Реакция, катализируемая ПДК, связывает между co6oй такие метаболические пути, как гликолиз (распад глюкозы), глюконеогенез (синтез глюкозы), синтез жирных кислот, окисление жирных кислот и цикл Кребса. Можно сказать, что реакции, катализируемые ПДК, представляют собой большой биохимический перекресток.
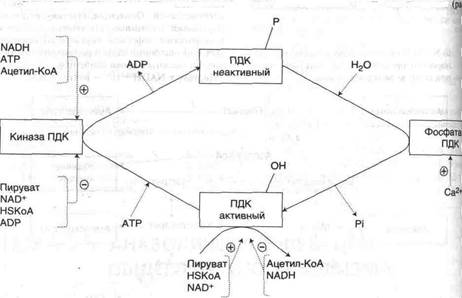
Рис. 9. Регуляция пируватдегидрогеназного комплекса
В составе ПДК содержатся 2 регуляторные субъединицы: киназа и фосфатаза. Киназа фосфорилирует ПДК и переводит его в неактивную форму, фосфатаза отщепляет фосфорный остаток от ПДК и переводит его в активную форму. Киназа ПДК аллостерически активируется АТФ, НАДН и ацетил-КоА, а ингибируется пируватом, АДФ, НАД, HSKoA, Са2+ (рис.10).
Киназа ПДК аллостерически активируется НАДН, ацетил-КоА и АТФ, следовательно, при их накоплении прекращается дальнейшее превращение пирувата в ацетил-КоА. Такая ситуация создается, например,в печени при голодании: из жировых депо в печень поступают жирные кислоты, в митохондриях в результате специфического пути их катаболизма накапливается большое количество ацетил-КоА и НАДН. Пируват при этом не окисляется и может быть использован для синтеза глюкозы (глюконеогенеза).
Киназа ПДК аллостерически иигибируется пируватом, AДФ, HSKoA, Са2+. В абсорбционный период глюкоза поступает в клетки и распадается с образованием пирувата. Высокая концентрация пирувата действует на ПДК двумя способами:
• поддерживает ПДК в нефосфорилированной активной форме, так как это наиболее сильный ингибитор киназы ПДК;
• аллостерически активирует нефосфорилированную активную форму ПДК, действуя согласованно с другими активаторами - субстратами реакций - НАД+ и HSKoA. В результате создаются условия для образования ацетил-КоА из глюкозы. Ацетил-КоА может окисляться в ЦТК; в печени и жировой ткани часть ацетил-КоА используется для синтеза жирных кислот.
Регуляция ионами Ca2+ особенно важна в мышцах. Потенциал действия увеличивает концентрацию Ca2+ в митохондриях, что одновременно ингибирует киназу и активирует фосфатазу; это быстро переводит ПДК в активную нефосфорилированную форму. Одновременно Са2+ активирует регуляторные ферменты ЦТК, и ацетил-КоА быстро окисляется, обеспечивая синтез АТР для работы мышц.
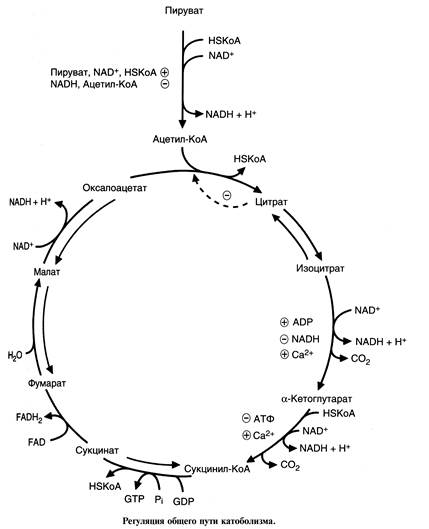
Рис. 10
В адипоцитах инсулин, действуя через мембранные рецепторы, приводит к увеличению концентрации Ca2'1' в митохондриях, что активирует фосфатазу ПДК и переводит его в активное нефосфорилированное состояние. В результате создаются условия для превращений: пируват ® ацетил-КоА ® жирные кислоты ® жиры, т.е. из продуктов распада глюкозы синтезируются жиры - основная форма запасания энергии в организме.
Регуляция ОПК дает возможность переключать метаболические пути, например в абсорбционный период продукты катаболизма глюкозы в печени используются для синтеза жиров, окисление жирных кислот в печени при голодании делает возможным использование пирувата для синтеза глюкозы.
Дата добавления: 2015-09-18; просмотров: 4633;