Генетический контроль функции клетки. Синтез белка.
Все клеточные функции осуществляются специфическими белками — ферментами. Поэтому основной механизм регуляции внутриклеточных процессов связан с влиянием на эти функции указанных белков. Эта регуляция осуществляется через усиление или ослабление синтеза ферментов на генетическом уровне, контролируемом ДНК, но может быть направлена и на изменение активности уже синтезированных ферментов, как стимулируя, так и тормозя ее. В обоих способах регуляции внутриклеточных процессов участвуют гормоны, медиаторы (вещества, выделяемые нервными окончаниями), а также продукты, синтезируемые в самой клетке. В последнем случае регуляция генетического контроля функции клеток осуществляется по принципу обратной связи.
ДНК передает генетический код клеткам-потомкам и воспроизводит этот код, т.е. управляет синтезом белков в клетке, определяя характер синтезируемых ферментов, структурных и секретируемых белков. Для выполнения первой задачи во время деления соматических клеток (митоза) две цепи, составляющие молекулу ДНК, разделяются, и каждая из них оказывается матрицей для синтеза новой цепи, подобной первой. Биосинтез новой цепи ДНК называется репликацией. Репликация ДНК катализируется ДНК-полимеразой и может продолжаться 8- 12 часов. Количество ДНК, представленное в каждой из двух дочерних клеток, равно ее количеству, содержащемуся в материнской клетке, т.к. в придачу к цепи ДНК, отделившейся во время митоза и переданной в дочернюю клетку, добавляется дополнительная, вновь синтезированная цепь ДНК. Эти клет-
ки называются диплоидными (2N). В случае деления половых клеток (мейоза) одна диплоидная клетка после двух быстро следующих друг за другом делений дает начало 4 клеткам, содержащим по одному набору хромосом, т.е. половину хромосомного материала по сравнению с содержащимся в соматических клетках. Эти клетки называются гаплоидными. Диплоидный набор восстанавливается объединением двух половых клеток, но половина ДНК происходит из половой клетки матери, а другая половина — из клетки отца.
Синтез белка и его регуляция.Управление синтезом белка включает в себя два этапа: 1) воспроизведение последовательности нук-леотидов, представленных в ДНК, в последовательностях РНК, называемое генетической транскрипцией, 2) использование информации РНК, для синтеза белков из аминокислот (трансляция).
| Рис. 1.10 Схема синтеза белка в клетке. |
Синтез белков организуется ДНК с различными типами РНК: информационной (иРНК), рибосомальной (рРНК), транспортной (тРНК). Первый тип — иРНК, синтезируется в ядре клетки. Ее синтез на одной из двух цепей ДНК катализируется РНК-полиме-разой (рис. 1.10). Синтезируемая иРНК повторяет последовательность нуклеотидов, составляющих генетический код ДНК. Генетический код представлен последовательностями триплетов оснований нуклеотидов, т.е. каждые три последовательных основания есть "слово"
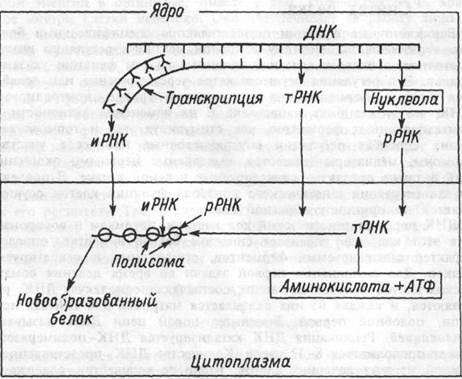
кода. Каждый триплет кодирует позицию одной аминокислоты. Отсюда триплеты иРНК определяют порядок включения аминокислот в молекулу белка во время ее синтеза в клетке. Например, два последовательных триплета (гуанин-гаунин-гаунин, ГГГ и гуанин-тимидин-тимидин, ГТТ) ответственны за размещение двух аминокислот — пролина и глютаминовой кислоты в молекуле белка. Кодирующий триплет иРНК называется кодоном. Следовательно, цепь кодонов, в свою очередь, составляет матрицу для синтеза аминокислотной цепи белка. Синтез иРНК предваряется активацией нук-леотидов, присоединением к каждому из них двух фосфатных радикалов, полученным от АТФ клетки, т.е. идет с потреблением энергии.
РНК-полимераза "узнает" участок, с которого начинается тран-крипция ДНК, так называемый промотор, присоединяется к нему, расплетает двойную спираль ДНК и, перемещаясь вдоль одной из ее цепей, вдоль составляющих ее структурных генов, образует нить РНК, подобную "списываемому" участку матрицы. Когда РНК-полимераза достигает конца копируемого участка {терминатора), на котором заканчивается информация о синтезируемом белке, синтез молекулы РНК прерывается, она отделяется от матрицы в нуклео-плазму, а двойная спираль ДНК вновь восстанавливается.
| Рис.1.11 Схема функции оперона. |
Описанная транскрипция генов ДНК происходит на участке, называемом опероном (рис.1.11). В его состав помимо промотора, структурных генов и терминатора входит оператор, расположенный за промотором, с которым взаимодействует регуляторный белок — репрессор. Репрессор, взаимодействуя с оператором, разрешает или тормозит движение РНК-полимеразы. Это объясняется тем, что репрессорный белок существует в двух различных формах, одна из которых, связываясь с оператором, тормозит транскрипцию, другая же этого эффекта не производит. Например, гемопротеин, связанный с молекулой кислорода, тормозит в клетках почек транскрип-
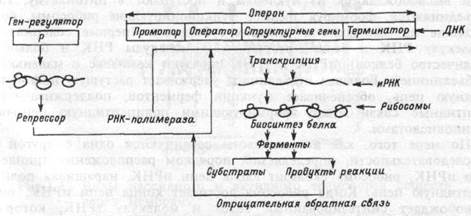
цию генов, ответственных за синтез гормона эритропоэтина. При гипоксии почек гемопротеин лишается молекулы О2, его сродство к оператору понижается и синтез эритропоэтина активируется (глава 6). Сродство репрессора к оператору может повышаться или понижаться под влиянием различных клеточных продуктов реакции, катализируемой ферментами, синтез которых контролируется оперо-ном. Так, увеличение количества синтезируемого вещества угнетает оперон по механизму отрицательной обратной связи, что позволяет клетке регулировать процесс транскрипции и количество синтезируемого белка. Обработанная ферментами нуклеоплазмы иРНК поступает через поры в ядре в цитоплазму и прикрепляется к рРНК.
Второй тип РНК — тРНК. В клетке много различных типов тРНК, но каждый из них комбинируется только с одной из 20 аминокислот, "узнает" кодон соответствующей аминокислоты на иРНК и транспортирует аминокислоту к этому месту. Таким образом, каждая тРНК является переносчиком специфической для нее аминокислоты к месту сборки белка — к полисомам. Аминокислоты вступают в синтез определенного белка после активации их молекулой АТФ, т.е. лишь активированная АТФ аминокислота соединяется с молекулой специфической тРНК. Специфический кодон в тРНК, который позволяет ей узнавать комплементарный кодон в иРНК — это также триплет нуклеотидных оснований и называется он антикодон. Во время формирования молекулы белка антикодо-новые основания соединяются водородными мостиками с основаниями кодона иРНК. Благодаря этому, аминокислоты выстраиваются одна за другой вдоль цепи иРНК, образуя соответствующую последовательность аминокислот в молекуле белка.
Третий тип РНК — рРНК. Ею образовано около 60% массы рибосом. Гены ДНК для формирования рРНК локализованы в пяти различных хромосомах, что связано с высокой потребностью клетки в данном типе РНК. Синтезированная рРНК накапливается в нук-леоле, где формируются изначальные субъединицы рибосом. Затем они высвобождаются из нуклеолы и поступают в цитоплазму, где объединяются, формируя зрелые, функционирующие рибосомы, состоящие из маленьких и больших субъединиц (первые содержат 1 молекулу РНК и белки, вторые — 3 молекулы РНК и большое количество белков). иРНК и тРНК образуют комплекс с маленькой субъединицей. Большая субъединица удерживает растущую полипептидную цепь, обеспечивает функции ферментов, поддерживающих пептидные связи между формирующими полипептидную цепочку аминокислотами.
По мере того, как аминокислоты соединяются одна с другой в последовательности, определяемой порядком расположения триплетов иРНК, рибосома скользит вдоль цепи иРНК, наращивая полипептидную цепь. Когда рибосома достигает конца цепи иРНК, она освобождает синтезированный белок и молекулу тРНК, которая вновь используется в трансляции. Клетками много раз может быть использована и молекула иРНК.
1.6. Трансмембранный пассивный транспорт.
У животных с замкнутой сосудистой системой внеклеточная жидкость условно разделяется на два компонента: интерстициальная жидкость и циркулирующая плазма крови. Интерстициальная жидкость представляет собой часть внеклеточной жидкости, которая расположена вне сосудистой системы и омывает клетки. Около 1/3 общей воды тела составляет внеклеточная жидкость, остальные 2/3 — жидкость внутриклеточная.
Концентрации электролитов и коллоидных веществ существенно отличаются в плазме, интерстициальной и внутриклеточной жидкостях. Наиболее выраженные различия состоят в относительно низком содержании белков-анионов в интерстициальной жидкости, в сравнении с внутриклеточной жидкостью и плазмой крови, и более высоких концентрациях натрия и хлора в интерстициальной, а калия во внутриклеточной жидкости.
Неодинаковый состав различных жидких сред тела в значительной степени обусловлен природой разделяющих их барьеров. Клеточные мембраны отделяют внутриклеточную от внеклеточной жидкости, стенки капилляров — интерстициальную жидкость от плазмы. Перенос веществ через эти барьеры может происходить пассивно за счет диффузии, фильтрации и осмоса, а также посредством активного транспорта.
Пассивный перенос веществ через клеточные мембраны не требует затраты энергии метаболизма. Активный транспорт осуществляется транспортными аденозинтрифосфатазами (АТФазами) и происходит за счет энергии гидролиза АТФ. Схематически основные виды транспорта веществ через мембрану клеток представлены на рис.1.12.
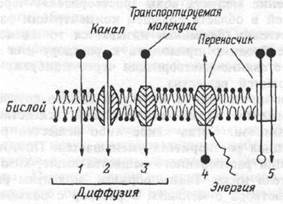
Рис.1.12 Виды пассивного и активного транспорта веществ через мембрану.
1,2 — простая диффузия через бислой и ионный канал,
3 — облегченная диффузия, 4 — первично-активный транспорт,
5 — вторично-активный транспорт.
Простая диффузия.Диффузия представляет собой процесс, при помощи которого газ или растворенные вещества распространяются и заполняют весь доступный объем.
Молекулы и ионы, растворенные в жидкости, находятся в хаотическом движении, сталкиваясь друг с другом, молекулами растворителя и клеточной мембраной. Столкновение молекулы или иона с мембраной может иметь двоякий исход: молекула либо "отскочит" от мембраны, либо пройдет через нее. Когда вероятность последнего события высока, то говорят, что мембрана проницаема для данного вещества.
Если концентрация вещества по обе стороны мембраны различна, возникает поток частиц, направленный из более концентрированного раствора в разбавленный. Диффузия происходит до тех пор, пока концентрация вещества по обе стороны мембраны не выравнивается. Через клеточную мембрану проходят как хорошо растворимые в воде {гидрофильные) вещества, так и гидрофобные, плохо или совсем в ней нерастворимые.
Гидрофобные, хорошо растворимые в жирах вещества, диффундируют благодаря растворению в липидах мембраны. Вода и вещества хорошо в ней растворимые проникают через временные дефекты углеводородной области мембраны, т.н. кинки, а также через поры, постоянно существующие гидрофильные участки мембраны.
В случае, когда клеточная мембрана непроницаема или плохо проницаема для растворенного вещества, но проницаема для воды, она подвергается действию осмотических сил. При более низкой концентрации вещества в клетке, чем в окружающей среде, клетка сжимается; если концентрация растворенного вещества в клетке выше, вода устремляется внутрь клетки.
Осмос— движение молекул воды (растворителя) через мембрану из области меньшей в область большей концентрации растворенного вещества. Осмотическим давлением называется то наименьшее давление, которое необходимо приложить к раствору для того, чтобы предотвратить перетекание растворителя через мембрану в раствор с большей концентрацией вещества.
Молекулы растворителя, как и молекулы любого другого вещества, приводятся в движение силой, возникающей вследствие разности химических потенциалов. Когда какое-либо вещество растворяется, химический потенциал растворителя уменьшается. Поэтому в области, где концентрация растворенного вещества выше, химический потенциал растворителя ниже. Таким образом, молекулы растворителя, перемещаясь из раствора с меньшей в раствор с большей концентрацией, движутся в термодинамическом смысле "вниз", "по градиенту".
Объем клеток в значительной степени регулируется количеством содержащейся в них воды. Клетка никогда не находится в состоянии полного равновесия с окружающей средой. Непрерывное движение молекул и ионов через плазматическую мембрану изменяет концентрацию веществ в клетке и, соответственно, осмотическое
давление ее содержимого. Если клетка секретирует какое-либо вещество, то для поддержания неизменной величины осмотического давления она должна либо выделять соответствующее количество воды, либо поглощать эквивалентное количество иного вещества. Поскольку среда, окружающая большинство клеток гипотонична, для клеток важно предотвратить поступление в них больших количеств воды. Поддержание же постоянства объема даже в изотонической среде требует расхода энергии, поэтому в клетке концентрация веществ неспособных к диффузии (белков, нуклеиновых кислот и т.д.) выше, чем в околоклеточной среде. Кроме того, в клетке постоянно накапливаются метаболиты, что нарушает осмотическое равновесие. Необходимость расходования энергии для поддержания постоянства объема легко доказывается в экспериментах с охлаждением или ингибиторами метаболизма. В таких условиях клетки быстро набухают.
Для решения "осмотической проблемы" клетки используют два способа: они откачивают в интерстиций компоненты своего содержимого или поступающую в них воду. В большинстве случаев клетки используют первую возможность — откачку веществ, чаше ионов, используя для этого натриевый насос (см.ниже).
В целом объем клеток, не имеющих жестких стенок, определяется тремя факторами: а) количеством содержащихся в них и неспособных к проникновению через мембрану веществ; б) концентрацией в интерстиций соединений, способных проходить через мембрану; в) соотношением скоростей проникновения и откачки веществ из клетки.
Большую роль в регуляции водного баланса между клеткой и окружающей средой играет эластичность плазматической мембраны, создающей гидростатическое давление, препятствующее поступлению воды в клетку. При наличии разности гидростатических давлений в двух областях среды вода может фильтроваться через поры барьера, разделяющего эти области.
Явления фильтрации лежат в основе многих физиологических процессов, таких, например, как образование первичной мочи в нефроне, обмен воды между кровью и тканевой жидкостью в капиллярах.
Диффузия ионов.Диффузия ионов происходит, в основном, через специализированные белковые структуры мембраны — ионные каналы, когда они находятся в открытом состоянии. В зависимости от вида ткани клетки могут иметь различный набор ионных каналов. Различают натриевые, калиевые, кальциевые, натрий-кальциевые и хлорные каналы. Перенос ионов по каналам имеет ряд особенностей, отличающих его от простой диффузии. В наибольшей степени это касается кальциевых каналов.
Ионные каналы могут находиться в открытом, закрытом и инак-тивированном состояниях. Переход канала из одного состояния в другое управляется или изменением электрической разности потенциалов на мембране, или взаимодействием физиологически активных веществ с рецепторами. Соответственно, ионные каналы подразделяют на потенциал-зависимые и рецептор-управляемые. Избира-
39
тельная проницаемость ионного канала для конкретного иона определяется наличием специальных селективных фильтров в его устье.
Облегченная диффузия.Через биологические мембраны кроме воды и ионов путем простой диффузии проникают многие вещества (от этанола до сложных лекарственных препаратов). В то же время даже сранительно небольшие полярные молекулы, например, гликоли, моносахариды и аминокислоты практически не проникают через мембрану большинства клеток за счет простой диффузии. Их перенос осуществляется путем облегченной диффузии. Облегченной называется диффузия вещества по градиенту его концентрации, которая осуществляется при участии особых белковых молекул-переносчиков.
Транспорт Na+, K+, Сl-, Li+, Ca2+, НСО3- и Н+ могут также осуществлять специфические переносчики. Характерными чертами этого вида мембранного транспорта являются высокая по сравнению с простой диффузией скорость переноса вещества, зависимость от строения его молекул, насыщаемость, конкуренция и чувствительность к специфическим ингибиторам — соединениям, угнетающим облегченную диффузию.
Все перечисленные черты облегченной диффузии являются результатом специфичности белков-переносчиков и ограниченным их количеством в мембране. При достижении определенной концентрации переносимого вещества, когда все переносчики заняты транспортируемыми молекулами или ионами, дальнейшее ее увеличение не приведет к возрастанию числа переносимых частиц — явление насыщения. Вещества, сходные по строению молекул и транспортируемые одним и тем же переносчиком, будут конкурировать за переносчик — явление конкуренции.
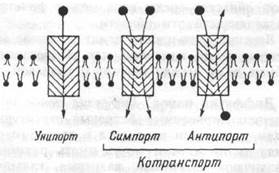
|
| Рис. 1.13 Классификация способов переноса через мембрану. |
Различают несколько видов транспорта веществ посредством облегченной диффузии (рис. 1.13): уни-порт, когда молекулы или ионы переносятся через мебрану независимо от наличия или переноса других соединений (транспорт глюкозы, аминокислот через базальную мембрану эпителиоцитов); симпорт, при котором их перенос осуществляется одновременно и однона-правленно с другими соединениями (натрий- зависимый транспорт Сахаров и аминокислот Na+ K+, 2Cl- и котран-спорт); антипорт — (транспорт вещества обусловлен одновременным и противоложно направленным транспортом другого соединения или иона (Na+/Ca2+, Na+/H+ Сl-/НСО3 -— обмены). Симпорт и
40
антипорт — это виды котранспорта, при которых скорость переноса контролируется всеми участниками транспортного процесса.
Природа белков-переносчиков неизвестна. По принципу действия они делятся на два типа. Переносчики первого типа совершают челночные движения через мембрану, а второго — встраиваются в мембрану, образуя канал. Промоделировать их действие можно с помощью антибиотиков-ионофоров, переносчиком щелочных металлов. Так, один из них — (валиномицин) — действует как истинный переносчик, переправляющий калий через мембрану. Молекулы же грамицидина А, другого ионофора, встаиваются в мембрану друг за другом, формируя "канал" для ионов натрия.
Большинство клеток обладают системой облегченной диффузии. Однако перечень метаболитов, переносимых с помощью такого механизма, довольно ограничен. В основном, это сахара, аминокислоты и некоторые ионы. Соединения, являющиеся промежуточными продуктами обмена (фосфорилированные сахара, продукты метаболизма аминокислот, макроэрги), не транспортируются с помощью этой системы. Таким образом, облегченная диффузия служит для переноса тех молекул, которые клетка получает из окружающей среды. Исключением является транспорт органических молекул через эпителий, который будет рассмотрен отдельно.
Активный транспорт.Транспорт веществ из среды с низкой концентрацией в среду с более высокой концентрацией не может быть объяснен движением по градиенту, т.е. диффузией. Этот процесс осуществляется за счет энергии гидролиза АТФ или энергии, обусловленной градиентом концентрации каких-либо ионов, чаще всего натрия. В случае, если источником энергии для активного транспорта веществ является гидролиз АТФ, а не перемещение через мембрану каких-то других молекул или ионов, транспорт называется первично активным.
Первично-активный перенос осуществляется транспортными АТФа-зами, которые получили название ионных насосов. В клетках животных наиболее распространена Na+ ,K+ — АТФаза (натриевый насос), представляющая собой интегральный белок плазматической мембраны и Са2+ — АТФазы, содержащиеся в плазматической мембране сарко-(эндо)-плазматического ретикулума. Все три белка обладают общим свойством — способностью фосфорилироваться и образовывать промежуточную фосфорилированную форму фермента. В фосфорилиро-ванном состоянии фермент может находиться в двух конформациях, которые принято обозначать Е1 и Е2. Конформация фермента — это способ пространственной ориентации (укладки) полипептидной цепи его молекулы. Две указанные конформации фермента характеризуются различным сродством к переносимым ионам, т.е. различной способностью связывать транспортируемые ионы.
Na+/K+- АТФаза обеспечивает сопряженный активный транспорт Na+ из клетки и К+ в цитоплазму. В молекуле Na+/K+- АТФазы имеется особая область (участок), в которой происходит связывание ионов Na и К. При конформации фермента E1 эта область обращена внутрь
41
клетки и обладает большим сродством к Na+, а при конформации Е2 — наружу и имеет высокое сродство к К+. В присутствии АТФ и Na+ в цитоплазме запускается фосфорилирование фермента и происходит присоединение 3-х ионов натрия к области связывания ионов в молекуле Na+/K+-АТФазы. В результате происходит такое изменение конформации фермента, при котором участок молекулы, присоединивший 3 иона натрия, оказывается на наружной стороне мембраны и его сродство к ионам натрия уменьшается (переход в форму Е2). Уменьшение сродства Na, К-АТФазы к Na+ приводит к освобождению этих ионов во внеклеточную жидкость.
В новой конформации фермента (Е2) его область связывания обладает высоким сродством к К+. Связывание 2-х ионов калия ведет к дефосфорилированию фермента и второму изменению конформации молекулы — переход в Е,. В конформации E1 область связывания ионов в молекуле Na+, К+ — АТФазы вновь обращена внутрь и имеет высокое сродство к Na+ и низкое к К+. Ионы калия освобождаются в цитоплазму и цикл работы фермента повторяется.
Таким образом, соотношение числа переносимых за один цикл работы фермента ионов натрия и калия и, соответственно, электрических зарядов равно 3/2. Следовательно, этот ионный насос является электрогенным — при его работе возникает чистый поток положительных зарядов из клетки — выходящий ток.
Активный транспорт Са2+ осуществляется Са2+-АТФазой, для которой также характерно циклическое изменение сродства к переносимому иону. Например, в скелетной мышце имеется сложная сеть трубочек и пузырьков — саркоплазматический ретикулум. Его основная функция — регуляция концентрации Са2+ в цитоплазме. Низкая концентрация Са2+ в цитоплазме в покоящейся мышце поддерживается благодаря работе Са2+-АТФазы мембраны саркоплазма-тического ретикулума. Цикл превращения этого фермента в процессе транспорта Са2+ из цитоплазмы, где его концентрация низкая (менее 10-7 М), в трубочки и пузырьки саркоплазматического ретикулума, где его концентрация высокая (10-3 — 10-2 М) состоит в следующем.
Когда в цитоплазме скелетного мышечного волокна присутствует АТФ и Са2+ происходит фосфорилирование Са2+-АТФазы и присоединение Са2+ к особой области фермента, которая называется кальций-связывающим участком. Этот участок молекулы Са2+-АТФазы в конформации Е, обращен в цитоплазму мышечного волокна. Фосфорилирование фермента и связывание Са2+ ведет к изменению конформации молекулы, в результате которого кальций-связывающий участок оказывается уже на стороне мембраны, обращенной в просвет саркоплазматического ретикулума. В новой конформации (Е2) фермент обладает меньшим сродством к Са2+, поэтому Са2+ отщепляется от него и поступает во внутриретикулярное пространство. Следующая стадия превращения фермента — дефосфорилирование и второе изменение конформации молекулы, при котором его сродство к Са2+ вновь увеличивается и кальций-связывающий участок оказывается на обращенной в цитоплазму стороне мембраны сарко-
42
плазматического ретикулума. Для осуществления этой стадии превращения Са2+-АТФазы необходимо присутствие в саркоплазмати-ческом ретикулуме ионов магния. В последующем цикл работы фермента повторяется.
Вторичный активный транспорт.Вторичным активным транспортом называется перенос через мембрану вещества против градиента его концентрации за счет энергии градиента концентрации другого вещества, создаваемого в процессе активного транспорта. В клетках животных основным источником энергии для вторичного активного транспорта служит энергия градиента концентрации ионов натрия, который создается за счет работы Na+/K+ - АТФазы. Например, мембрана клеток слизистой оболочки тонкого кишечника содержит белок, осуществляющий перенос (симпорт) глюкозы и Na+ в эпителиоциты. Транспорт глюкозы осуществляется лишь в том случае, если Na+, одновременно с глюкозой связываясь с указанным белком, переносится по электрохимическому градиенту. Электрохимический градиент для Na+ поддерживается активным транспортом этих катионов из клетки.
В головном мозге работа Na+-насоса сопряжена с обратным поглощением (реабсорбцией) медиаторов — физиологически активных веществ, которые выделяются из нервных окончаний при действии возбуждающих факторов.
В кардиомиоцитах и гладкомышечных клетках с функционированием Na+, K+-АТФазы связан транспорт Са2+ через плазматическую мембрану, благодаря присутствию в мембране клеток белка, осуществляющего противотранспорт (антипорт) Na+ и Са2+. Ионы кальция переносятся чере мембрану клеток в обмен на ионы натрия и за счет энергии концентрационного градиента ионов натрия.
В клетках обнаружен белок, обменивающий внеклеточные ионы натрия на внутриклеточные протоны — Na+/H+ — обменник. Этот переносчик играет важную роль в поддержании постоянства внутриклеточного рН. Скорость, с которой осуществляется Na+/Ca2+ и Na+/H+ — обмен, пропорциональна электрохимическому градиенту Na+ через мембрану. При уменьшении внеклеточной концентрации Na+ ингибировании Na+ , K+-АТФазы сердечными гликозидами или в бескалиевой среде внутриклеточная концентрация кальция и протонов увеличена. Это увеличение внутриклеточной концентрации Са2+ при ингибировании Na+, K+-АТФазы лежит в основе применения в клинической практике сердечных гликозидов для усиления сердечных сокращений.
Дата добавления: 2015-09-11; просмотров: 2049;