Эндонуклеазы рестрикции типа II
В системах рестрикции-модификации II-го типа нуклеазная и метилтрансферазная активности в большинстве случаев обеспечиваются независимыми белками. Рестрицирующие ферменты, входящие в системы этого типа, включают 11 подгрупп, которые отличаются, в основном, по структуре участка узнавания для фермента, а также по расположению позиции расщепления обеих нитей ДНК относительно узнаваемого сайта. Эндонуклеазы II-го типа разрезают ДНК в строго определенных позициях внутри или около сайта узнавания, в результате концы ДНК в месте разрыва имеют 3’-гидроксильные и 5’-фосфатные группы, способные к восстановлению фосфодиэфирных связей, т.е. к восстановлению полинуклеотидной цепочки в обеих нитях ДНК. Ввиду данных особенностей, эндонуклеазы этого типа особенно широко используются в генетической инженерии для получения рекомбинантных ДНК.
Для функционирования ферментов типа II не требуется АТФ или другие источники энергии. В системах рестрикции-модификации из разных источников активные нуклеазы могут существовать в виде мономера, гомодимера или гомотетрамера.
Большинство рестрицирующих эндонуклеаз класса II узнают последовательности, содержащие от 4 до 6 нуклеотидных пар, поэтому эти ферменты делят на мелко- и крупнощепящие. Мелкощепящие эндонуклеазы узнают тетрануклеотид и вносят в молекулы гораздо больше разрывов, чем крупнощепящие, узнающие последовательность из шести нуклеотидных пар. Это связано с тем, что вероятность встречаемости определенной последовательности из четырех нуклеотидов гораздо выше, чем последовательности из шести нуклеотидов. Например, в ДНК бактериофага Т7, состоящей из 40000 пар оснований, отсутствует последовательность, представляющая собой гексануклеотид GAATTC, который узнает фермент EcoRI (из Escherichia coli). Например, к мелкощепящим относятся эндонуклеазы HpaII (из Haemophilus parainfluenzae) и Alu (из Arthrobacter luteus), к крупнощепящим - EcoRI и HindIII (из Haemophilus influenzae). Если предположить, что участки узнавания эндонуклеаз рестрикции распределены вдоль цепи ДНК случайно, то мишень для ферментов, узнающих последовательность из четырех нуклеотидов, должна встречаться в среднем 1 раз через каждые 256 пар оснований, а для ферментов, узнающих шесть нуклеотидов, - через 4096 пар оснований.
Последовательности нуклеотидов двуцепочечной ДНК, узнаваемые эндонуклеазами рестрикции II типа (сайты рестрикции) часто представляют собой палиндромы. Точки узнавания рестриктазами симметричны относительно поворота на 180оС, то есть последовательность нуклеотидов слева направо в одной нити ДНК такая же, как справа налево в другой (рис. 2).Палиндромные участки узнавания и разрезания могут быть представлены также пятью парами, где средний нуклеотид может быть любым из четырех нуклеотидов (HinfI),
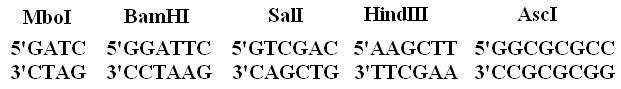
Рис. 2. Примеры палиндромных участков узнавания ДНК для эндонуклеаз рестрикции.
Расщепление происходит в фиксированном положении внутри сайта рестрикции или непосредственно вблизи него. При этом различные фрагменты вызывают образование как липких, так и тупых концов на фрагментах ДНК.
Эндонуклеазы рестрикции могут узнавать непалиндромные участки и разрезать ДНК на некотором расстоянии от них. В таких случаях образуются одноцепочечные выступащие концы длиной в один нуклеотид
• MboII 5’– GGTGANNNNNNNN↓–
• CCACTNNNNNNN↑ - 5’
Полностью метилированный сайт не подвержен ни рестрикции, ни модификации. Полуметилированный сайт не узнается ферментом рестрикции, но может быть превращен с помощью метилтрансферазы в полностью метилированный. У бактерий метилирование, как правило, связано с сохранением имеющегося состояния модификации. Репликация полностью метилированной ДНК ведет к образованию полуметилированной ДНК. Вероятно, узнавание полуметилированных сайтов представляет собой обычный этап функционирования метилтрансферазы in vivo. Неметилированный сайт-мишень представляет собой субстрат либо для рестрикции, либо для модификации in vitro. В клетке немодифицированная ДНК с большей вероятностью будет рестрицирована. Реакция разрезания осуществляется в две ступени. Сначала происходит гидролиз одной цепи ДНК, а затем – другой.
Среди пятисот рестрицирующих ферментов, выделенных из различных микроорганизмов, встречаются такие, которые узнают на ДНК одни и те же последовательности. Такие пары или группы называют изошизомерами. Первый выделенный фермент для узнавания и специфического разрезания заданной последовательности, называют прототипом, а все остальные подобные рестриктазы называют изошизомерами. Различают истинную изошизомерию: SphI (CGTAC↓G) и BbuI (CGTAC↓G), когда ферменты узнают и гидролизуют одну и ту же последовательность нуклеотидов ДНК в одних и тех же точках, и ложную,: XmaI (C↓CCGGG) и SmaI (CCC↓GGG), когда ферменты, узнавая один и тот же сайт на ДНК, производят разрывы в разных точках в пределах того же сайта. Совсем недавно предложено такие ферменты, как XmaIи SmaI, называть гетерошизомерами. Рестриктазы, разпознающие совершенно разные последовательности, но образующие одинаковые концы, называют изокаудомерами: MboI (↓GATC) и BamHI (G↓GATCC).
Дата добавления: 2015-07-14; просмотров: 1370;