Биологическое окисление
При биологическом окислении от органической молекулы под действием соответствующего фермента отщепляются два атома водорода. В ряде случаев при этом между ферментами и окисленной молекулой образуется неустойчивая, богатая энергией (макроэнергетическая) связь. Она используется для образования АТФ — "конечной цели" большинства процессов биологического окисления. А два отнятых атома водорода оказываются в результате реакции связанными с коферментом НАД (никотинамидадениндинуклеотидом) или с НАДФ (никотинамидаденин-динуклеотидфосфатом).
Дальнейшая судьба водорода может быть различной. При анаэробном окислении он переносится на некоторые органические молекулы. При аэробном окислениии водород передаётся на кислород с образованием воды. Основная часть цепи переноса водорода расположена в мембранах митохондрий. При этом из АДФ и неорганического фосфата образуется АТФ.
Надо отметить, что аэробное окисление намного эффективнее анаэробного. В первом случае из 1 молекулы глюкозы образуется 2 молекулы АТФ, а во втором — 36, где глюкоза "сжигается" до CO2 и воды. Это и объясняет широкое распространение и бурную эволюцию аэробных организмов.
Основной источник энергии в клетке - окисление субстратов кислородом воздуха. Этот процесс осуществляется тремя путями: присоединением кислорода к атому углерода, отщеплением водорода или потерей электрона. В клетках окисление протекает в форме последовательного переноса водорода и электронов от субстрата к кислороду. Кислород играет в этом случае роль восстанавливающегося соединения (окислителя). Окислительные реакции протекают с высвобождением энергии. Для биологических реакций характерны сравнительно небольшие изменения энергии. Это достигается за счет дробления процесса окисления на ряд промежуточных стадий, что позволяет запасать ее небольшими порциями в виде макроэргических соединений (АТФ). Восстановление атома кислорода при взаимодействии с парой протонов и электронов приводит к образованию молекулы воды.
Тканевое дыхание. Это процесс потребление клетками тканей организма кислорода, который участвует в биологическом окислении. Такой вид окисления называют аэробным окислением. Если конечным акцептором в цепи переноса водорода выступает не кислород, а другие вещества (например пировиноградная кислота), то такой тип окисления называют анаэробным.
Т.о. биологическое окисление - это дегидрирование субстрата с помощью промежуточных переносчиков водорода и его конечного акцептора.
Дыхательная цепь (ферменты тканевого дыхания) - это переносчики протонов и электронов от окисляемого субстрата на кислород. Окислитель - это соединение, способное принимать электроны. Такая способность количественно характеризуется окислительно-восстановительным потенциалом по отношению к стандартному водородному электроду, рН которого равен 7,0. Чем меньше потенциал соединения, тем сильнее его восстанавливающие свойства и наоборот.
Т. о. любое соединение может отдавать электроны только соединению с более высоким окислительно-восстановительным потенциалом. В дыхательной цепи каждое последующее звено имеет более высокий потенциал, чем предыдущее.
Дыхательная цепь состоит из:
1 НАД - зависимой дегидрогеназы;
2 ФАД- зависимой дегидрогеназы;
3 Убихинона (КоQ);
4 Цитохрмов b, c, a+a3 .
НАД-зависимые дегидрогеназы. В качестве кофермента содержат НАД и НАДФ. Пиридиновое кольцо никотинамида способно присоединять электроны и протоны водорода.
ФАД и ФМН-зависимые дегидрогеназы содержат в качестве кофермента фосфорный эфир витамина В2 (ФАД).
Убихинон (КоQ) отнимает водород у флавопротеидов и превращается при этом в гидрохинон.
Цитохромы - белки хромопротеиды, способные присоединять электроны, благодаря наличию в своем составе в качестве простетических групп железопорфиринов. Они принимают электрон от вещества, являющегося немного боле сильным восстановителем, и передают его более сильному окислителю. Атом железа связан с атомом азота имидазольного кольца аминоксилоты гистидина с одной стороны от плоскости порфиринового цикла, а с другой стороны с атомом серы метионина. Поэтому потенциальная способность атома железа в цитохромах к связыванию кислорода подавлена.
В цитохроме спорфириновая плоскость ковалентно связана с белком через два остатка цистеина, а в цитохромах b и а, она ковалентно не связанос белком.
В цитохроме а+а3(цитохромоксидазе) вместо протопорфирина содержатся порфирин А, который отличатся рядом структурных особенностей. Пятое координационное положение железа занято аминогруппой, принадлежащей остатку аминосахара, входящего в состав самого белка.
В отличие от гема гемолгобина атом железа в цитохромах может обратимо переходить из двух в трехвалентное состояние, это обеспечивает транспорт электронов.
Механизм работы электронтранспортной цепи. Наружная мембрана митохондрий проницаема для большинства мелких молекул и ионов, внутренняя почти для всех ионов (кроме протонов Н) и для большинства незаряженных молекул.
Процессы окисления и образования АТФ из АДФ и фосфорной кислоты, т.е. фосфорилирования протекают в митохондриях на внутренней мембране - кристах. Такая молекула содержит в себе три макроэргических связи. Макроэргической или богатой энергией называют химическую связь, при разрыве которой высвобождается более 4 ккал/моль. При гидролитическом расщеплении АТФ до АДФ и фосфорной кислоты высвобождается 7,3 ккал/моль. Ровно столько же тратится для образования АТФ из АДФ и остатка фосфорной кислоты и это один из основных путей запасания энергии в организме.
В процессе транспорта электронов по дыхательной цепи высвобождается энергия, которая тратится на присоединение остатка фосфорной кислоты к АДФ с образованием одной молекулы АТФ и одной молекулы воды. В процессе переноса одной пары электронов по дыхательной цепи высвобождается и запасается в виде трех молекул АТФ 21,3 ккал/моль. Это составляет около 40 % высвободившейся при электронном транспорте энергии.
Такой способ запасания энергии в клетке называется окислительным фосфорилированием или сопряженным фосфорилированием.
Скорость окислительного фосфорилирования зависит в первую очередь от содержания АТФ, чем быстрее она расходуется, тем больше накапливается АДФ, тем больше потребность в энергии и следовательно активнее идет процесс окислительного фосфорилирования. Регуляцию скорости окислительного фосфорилирования концентрацией в клетке АДФ называют дыхательным контролем.
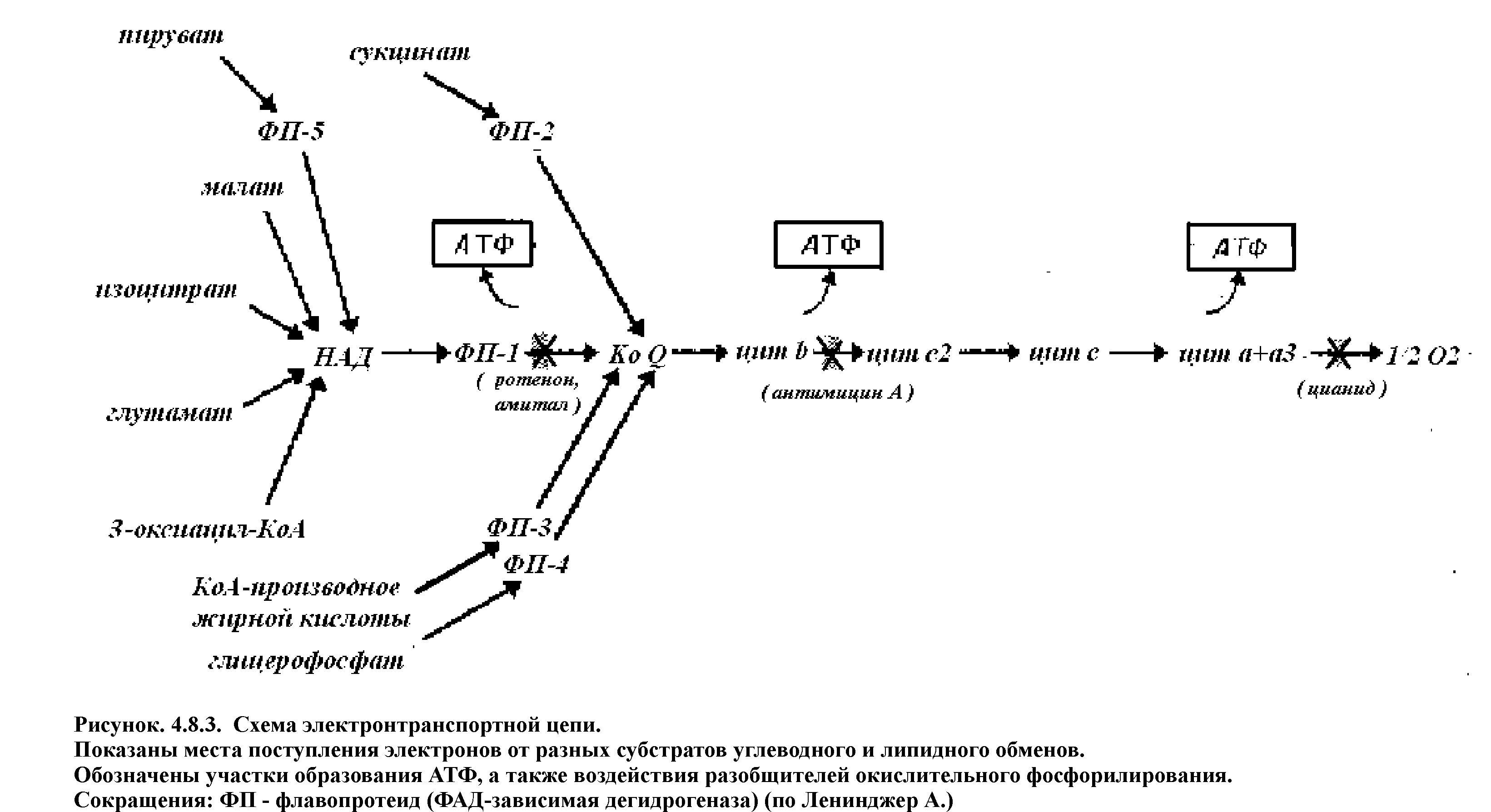
Рисунок 27-Дыхательный контроль
Дата добавления: 2015-08-11; просмотров: 3188;