Проводящие ткани
Проводящие ткани служат для передвижения по растению растворенных в воде питательных веществ. Они возникли как следствие приспособления растений к жизни на суше. В связи с жизнью в двух средах – почвенной и воздушной, возникли две проводящие ткани, по которым вещества передвигаются в двух направлениях. По ксилеме от корней к листьям поднимаются вещества почвенного питания – вода и растворенные в ней минеральные соли (восходящий, или транспирационный ток). По флоэме от листьев к корням передвигаются вещества, образовавшиеся в процессе фотосинтеза, главным образом сахароза (нисходящий ток). Так как эти вещества представляют собой продукты ассимиляции углекислого газа, транспорт веще ств по флоэме называют током ассимилятов.
Проводящие ткани образуют в теле растения непрерывную разветвленную систему, соединяющую все органы – от тончайших корешков до самых молодых побегов. Ксилема и флоэма представляют собой сложные ткани, в их состав входят разнородные элементы – проводящие, механические, запасающие, выделительные. Самыми важными являются проводящие элементы, именно они выполняют функцию проведения веществ.
Ксилема и флоэма формируются из одной и той же меристемы и, поэтому, в растении всегда располагаются рядом. Первичные проводящие ткани образуются из первичной латеральной меристемы – прокамбия, вторичные – из вторичной латеральной меристемы – камбия. Вторичные проводящие ткани имеют более сложное строение, чем первичные.
Ксилема (древесина) состоит из проводящих элементов – трахеид и сосудов (трахей), механических элементов - древесинных волокон (волокон либриформа) и элементов основной ткани - древесинной паренхимы.
Проводящие элементы ксилемы носят название трахеальных элементов. Различают два типа трахеальных элементов – трахеиды и членики сосудов (рис. 3.26 ).
Трахеида представляет собой сильно вытянутую в длину клетку с ненарушенными первичными стенками. Передвижение растворов происходит путем фильтрации через окаймленные поры. Сосуд состоит из многих клеток, называемых члениками сосуда. Членики расположены друг над другом, образуя трубочку. Между соседними члениками одного и того же сосуда имеются сквозные отверстия – перфорации. По сосудам растворы передвигаются значительно легче, чем по трахеидам.
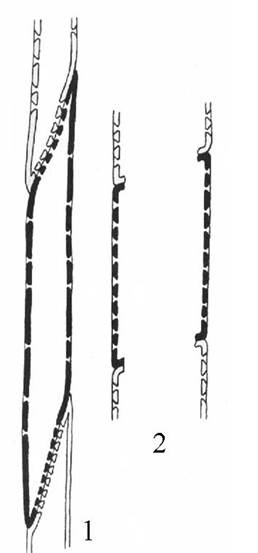
Рис. 3.26. Схема строения и сочетания трахеид (1) и члеников сосуда (2).
Трахеальные элементы в зрелом, функционирующем состоянии – мертвые клетки, не имеющие протопластов. Сохранение протопластов затрудняло бы передвижение растворов.
Сосуды и трахеиды передают растворы не только в вертикальном, но и в горизонтальном направлении в соседние трахеальные элементы и в живые клетки. Боковые стенки трахеид и сосудов сохраняются тонкими на большей или меньшей площади. В то же время они имеют вторичные утолщения, придающие стенкам прочность. В зависимости от характера утолщений боковых стенок трахеальные элементы называются кольчатыми, спиральными, сетчатыми, лестничными и точечно-поровыми (рис. 3.27).
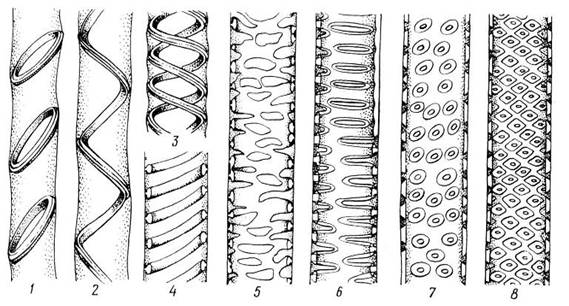
Рис. 3.27. Типы утолщения и поровости боковых стенок у трахеальных элементов : 1 – кольчатое, 2-4 – спиральные, 5 – сетчатое утолщения; 6 – лестничная, 7 – супротивная, 8 – очередная поровость.
Вторичные кольчатые и спиральные утолщения прикрепляются к тонкой первичной стенке посредством узкого выступа. При сближении утолщений и образовании между ними перемычек возникает сетчатое утолщение, переходящее в окаймленные поры. Эту серию (рис. 3.27 ) можно рассматривать как морфогенетический, эволюционный ряд.
Вторичные утолщения клеточных стенок трахеальных элементов одревесневают (пропитываются лигнином), что придает им дополнительную прочность, но ограничивает возможности роста в длину. Поэтому в онтогенезе органа сначала появляются еще способные растягиваться кольчатые и спиральные элементы, не препятствующие росту органа в длину. Когда рост органа прекращается, возникают элементы, неспособные к продольному растяжению.
В процессе эволюции первыми появились трахеиды. Они найдены у первых примитивных наземных растений. Сосуды появились значительно позже путем преобразования трахеид. Сосудами обладают почти все покрытосеменные растения. Споровые и голосеменные растения, как правило, лишены сосудов и обладают только трахеидами. Лишь в виде редкого исключения сосуды встречены у таких споровых, как селагинелла, некоторых хвощей и папоротников, а также у немногих голосеменных (гнетовые). Однако у этих растений сосуды возникли независимо от сосудов покрытосеменных. Возникновение сосудов у покрытосеменных растений означало важное эволюционное достижение, так как облегчило проведение воды; покрытосеменные растения оказались более приспособленными к жизни на суше.
Древесинная паренхима и древесинные волокна выполняют запасающие и опорные функции соответственно.
Флоэма (луб) состоит из проводящих - ситовидных - элементов, сопровождающих клеток (клеток-спутниц), механических элементов – флоэмных (лубяных) волокон и элементов основной ткани – флоэмной (лубяной) паренхимы.
В отличие от трахеальных элементов проводящие элементы флоэмы и в зрелом состоянии остаются живыми, а их клеточные стенки – первичными, неодревесневшими. На стенках ситовидных элементов имеются группы мелких сквозных отверстий – ситовидные поля, через которые сообщаются протопласты соседних клеток и происходит транспорт веществ. Различают два типа ситовидных элементов – ситовидные клетки и членики ситовидных трубок.
Ситовидные клетки являются более примитивными, они присущи споровым и голосеменным растениям. Ситовидная клетка – это одна клетка, сильно вытянутая в длину, с заостренными концами. Ее ситовидные поля рассеяны по боковым стенкам. Кроме того, ситовидные клетки имеют и другие примитивные признаки: они лишены специализированных сопровождающих клеток и в зрелом состоянии содержат ядра.
У покрытосеменных растений транспорт ассимилятов осуществляют ситовидные трубки(рис. 3.28 ). Они состоят из многих отдельных клеток – члеников, расположенных один над другим. Ситовидные поля двух соседних члеников образуют ситовидную пластинку. Ситовидные пластинки имеют более совершенное строение, чем ситовидные поля (перфорации крупнее и их больше).
В члениках ситовидных трубок в зрелом состоянии отсутствуют ядра, однако они остаются живыми и деятельно проводят вещества. Важная роль в проведении ассимилятов по ситовидным трубкам принадлежит сопровождающим клеткам (клеткам-спутницам). Каждый членик ситовидной трубки и его сопровождающая клетка (или две-три клетки в случае дополнительного деления) возникают одновременно из одной меристематической клетки. Клетки–спутницы имеют ядра и цитоплазму с многочисленными митохондриями; в них происходит интенсивный обмен веществ. Между ситовидными трубками и прилегающими к ним сопровождающими клетками имеются многочисленные цитоплазматические связи. Считается, что клетки-спутницы вместе с члениками ситовидных трубок составляют единую физиологическую систему, осуществляющую ток ассимилятов.
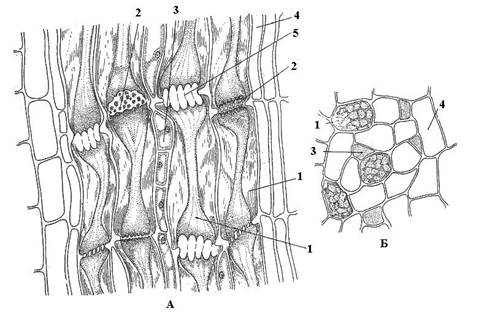
Рис. 3.28. Флоэма стебля тыквы на продольном (А) и поперечном (Б) срезе : 1 – членик ситовидной трубки; 2 – ситовидная пластинка; 3 – сопровождающая клетка; 4 – лубяная (флоэмная) паренхима; 5 – закупоренная ситовидная пластинка.
Длительность функционирования ситовидных трубок невелика. У однолетников и в надземных побегах многолетних трав – не более одного вегетационного периода, у кустарников и деревьев – не более трех-четырех лет. При отмирании живого содержимого ситовидной трубки, отмирает и клетка-спутница.
Лубяная паренхима состоит из живых тонкостенных клеток. В ее клетках часто накапливаются запасные вещества, а также смолы, танниды и др. Лубяные волокна играют опорную роль. Они присутствуют не у всех растений.
В теле растения ксилема и флоэма расположены рядом, образуя или слои, или обособленные тяжи, которые называют проводящимипучками. Различают несколько типов проводящих пучков (рис. 3.29 ).
Закрытые пучки состоят только из первичных проводящих тканей, они не имеют камбия и далее не утолщаются. Закрытые пучки характерны для споровых и однодольных растений. Открытые пучки имеют камбий и способны к вторичному утолщению. Они характерны для голосеменных и двудольных растений.
В зависимости от взаимного расположения флоэмы и ксилемы в пучке различают следующие типы. Наиболее обычны коллатеральные пучки, в которых флоэма лежит по одну сторону от ксилемы. Коллатеральные пучки могут быть открытыми (стебли двудольных и голосеменных растений) и закрытыми (стебли однодольных растений). Если с внутренней стороны от ксилемы располагается дополнительно тяж флоэмы, такой пучок называется биколлатеральным. Биколлатеральные пучки могут быть только открытыми, они характерны для некоторых семейств двудольных растений (тыквенные, пасленовые и др.).
Встречаются также концентрические пучки, в которых одна проводящая ткань окружает другую. Они могут быть только закрытыми. Если в центре пучка находится флоэма, а ксилема ее окружает, пучок называется центрофлоэмным, или амфивазальным. Такие пучки часто встречаются в стеблях и корневищах однодольных растений. Если в центре пучка располагается ксилема, и ее окружает флоэма, пучок называется центроксилемным, или амфикрибральным. Центроксилемные пучки обычны у папоротников.
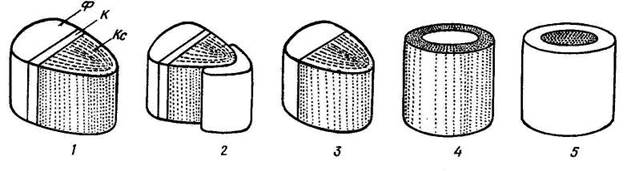
Рис. 3.29. Типы проводящих пучков : 1 – открытый коллатеральный; 2 – открытый биколлатеральный; 3 – закрытый коллатеральный; 4 – концентрический закрытый центрофлоэмный; 5 – концентрический закрытый центроксилемный; К – камбий; Кс – ксилема; Ф – флоэма.
Многие авторы выделяют радиальные пучки. Ксилема в таком пучке располагается в виде лучей от центра по радиусам, а флоэма – между лучами ксилемы. Радиальный пучок – характерный признак корня первичного строения.
Дата добавления: 2015-08-08; просмотров: 4791;