Такие ультраструктурные перестройки относитсяк обратимым пластическим реакциям одноклеточного организма.
Л. Н. Серавин (1969, 1978) предостерегает против механического использования применительно к простейшим широко распространенного термина «обучение». Безнервные одноклеточные животные, вероятно, обладают совершенно иными механизмами обучения, чем многоклеточные. Модификации поведения инфузорий могут носить совершенно иной характер. В качестве примера Л. Н. Серавин предлагает разработанную им модель модификации пищевого поведения у хишных инфузорий: путем изменения химических свойств пищи у простейших можно целенаправленно усилить пищевую реакцию за счет адекватной настройки их хеморецепторов на определенный индуктор пищевого поведения. Соответственно, при смене корма происходит перестройка хеморецеито-ров на индукторные вещества, выделяемые новой жертвой. Такая форма обучения носит избирательный характер и может сохраняться десятки дней.
Кишечнополостные,Среди многоклеточных самое низкое положение в эволюционной лестнице занимают ки-шечнополостные, у которых впервые появляется хотя и слабо дифференцированная, но организованная нервная система, построенная по диффузному типу.
Пресноводная гидра является излюбленным объектом для изучения с позиций сравнительной физиологии поведения. У гидры осуществление суммационного рефлекса одинаково хорошо достигается при любом порядке индифферентного и безусловного раздражений или только при одном из этих раздражений. Например, было показано, что у гидры при ограничении направленности движений тела поисковые движения продолжают осу-
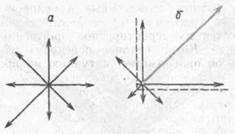
Рис. 8. Направленность поисковых движений стебельчатой гидры (по Л. Г Воронину. Б. Ф. Сергееву, 1979):
а — в норме (в течение часа); б — после часового ограничения ее движений слева и снизу; стрелки — направление и относительное число поисковых движений; прерывистой линией обозначен сектор размером 90°, в направлении которого во время тренировки могли осуществляться поисковые движения
ществляться преимущественно в направлении ранее разрешенного сектора еще длительное время после снятия ограничений (Б. Ф. Сергеев, 1979) (рис. 8).
Что касается привыкания, то новым его свойством, по сравнению с простейшими, является тренированность от опыта к опыту при интервалах между опытами, сравнимых с временем сохранения следа (Н. Л. Тушмалова, 1986).
Впервые в эволюции у червей начинается процесс централизации нервной системы и одновременный процесс начальной цефализации. Центральная нервная система построена по принци пу лестницы с двусторонней симметрией и разрастанием объема ганглиев, расположенных в головном конце тела. Однако у низших червей, типичным представителем которых являются плана рии, церебральный ганглий 'имеет черты весьма примитивной организации: в нем много мышечных клеток и элементов, выполняющих нсйросек-реторные функции. Примитивный уровень «условных рефлексов» коррелирует с достаточно низкой организацией головного ганглия планарий.
В то же время реакция сенсити-зации проявляется у планарий достаточно отчетливо, хотя и не обладает большой прочностью. Примитивные нестабильные «условные рефлексы» сходны по многим параметрам с сум-мационными рефлексами.
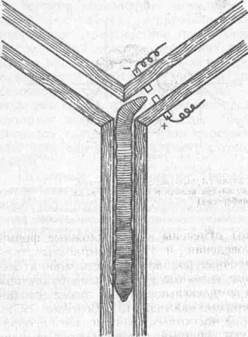
РИС.9 Установка для научения дождевого червя
(пo Э. К.,Фабри ,1976);
движение в правый рукав лабиринта сопровождаете» ударом электрического тока, что побуждает животное двигаться только и левый рукав лабиринта
Иное дело у высших (кольчатых) червей, куда относятся и обычные дождевые черви. Наряду с усложнением строения нервной системы, заключающемся в дальнейшем увеличении окологлоточного нервного кольца — предшественника головного мозга — расширяется разнообразие форм активного поведения этих животных. Большую группу поведенческих реакций, которые имеют генетическую основу, называют таксисами. Например, реакция дождевого червя на условия освещения состоит в том, что он уползает в сторону большего затемнения (отрицательный фототаксис).
«Условные рефлексы» характеризуются большим несовершенством. Так, дождевых червей обучали поворачивать в Т-образном лабиринте в левый рукав. Безусловным раздражением служил удар током в правом рукаве, а условным сам лабиринт, элементы которого воспринимались внешней ре-
цепцией (рис. 9). Условный рефлекс вырабатывался медленно, потребовалось около 400 сочетаний, по мере сочетаний увеличение числа правильных реакций не наступало. Удаление надглоточного ганглия не изменяло динамику обучения, что свидетельствует о незначительной цефализации у данных животных.
У морских кольчатых червей — ио-лихет возникают специальные ассоциативные центры «мозга» — грибовидные тела. У полихет показана возможность выработки стойких условных пищевых рефлексов, которые угасали при неподкреплении и самопроизвольно восстанавливались {Л. Г. Воронин, 1977).
Моллюски характеризуются типом организации нервной системы, получившим наименование разбросанно узловой, при котором проявляется как дальнейшая централизация нервных элементов в ограниченное число ганглиев, так и субординация с выделением в качестве ведущего органа управления поведением — зрительных долей «мозга» у головоногих моллюсков. Например, у осьминога (Д. Янг, 1964) зрительные доли выступают как высший интегра-тивный нервный аппарат, связанный со всеми остальными ганглиями. Соответственно и поведение этих «приматов моря» (И. Акимушкин, 1963) представляется чрезвычайно сложным, где врожденные комплексы поведения дополняются богатым индивидуальным опытом. Так, например, у осьминога легко и быстро вырабатывается условно-рефлекторное изменение окраски кожи при сочетании света с механическим кожным раздражением. Наряду с инструментальными пищедобыватель-ными условными рефлексами образуются и дифференцировочное торможение, и условный тормоз (см. гл. IV).
Более просто организованные брюхоногие моллюски являются излюбленным объектом для исследования некоторых сторон нейронных механизмов обучения (Э. Кендел, 1980; Е. Н. Соколов, 1981). Сравнительно простая нервная система позволяет изучать внутриклеточные механизмы привыкания, сен-ситизации, условных рефлексов. Напри-
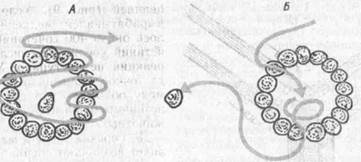
Рис. 10: Поведение роющей осы (по Э. К. Фабри, 1976):
после отлета оси кольцо из шишек вокруг норки (А) было перемешено в сторону (В). После возвращения оса ищет норку внутри кольца и находи! ее лишь после повторных облетов (Б) (опыт Н. Тинбергена)
мер, при неожиданном стимуле у пру* довика возникает рефлекс втягивания тела в раковину, которая накрывает голову, щупальца отводятся вниз и локомоция прекращается. При повторном воздействии механического стимула латентный период уменьшается, а амплитуда рефлекса увеличивается возникает сенситизация, которая может быть растянута на несколько дней. Оказалось, что облегчение может тесно взаимодействовать с привыканием этого же рефлекса. Скорее всего механизмы привыкания состоят в функциональном изменении синаптических контактов между сенсорным и двигательным нейронами.
Условные же рефлексы у моллюсков (аплизия, виноградная улитка) вырабатываются относительно легко и служат удобной моделью для изучения элементарных клеточных механизмов обучения и памяти (Г, Г. Аракелов, 1985; П. М. Балабан, 1985).
Другая систематическая группа животных, связанная происхождением с круглыми червями (аннелидами), представлена членистоногими, поведение которых наиболее тщательно изучалось на представителях ракообразных и особенно насекомых.
Низшим ракообразным (дафниям) свойственны лишь элементарные формы временных связей при сочетании света с низкой температурой и затемнения— с высокой. Избирательное реагирование животных сохранялось около одного часа.
У высших ракообразных (крабы, ра-
ки) отмечены весьма сложные формы поведения, и у них вырабатываются прочные положительные и отрицательные условные рефлексы на одиночные и комплексные сигналы, сложные лабиринтные навыки (Л. Г. Воронин, 1970).'
У насекомых головные ганглии образуют мощный головной мозг в виде сложно устроенного надглоточного узла, состоящего из трех отделов, передний из которых образует высший интс-гративный отдел, получивший название грибовидных тел. Продолжающаяся це-фализация, выраженная в большой концентрации нервных элементов головного ганглия, сопровождается значительным усложнением форм поведения.
Как и у головоногих моллюсков, у насекомых можно образовать истинные условные рефлексы спустя 15— 25 сочетаний, они могут подвергаться угашению и самовосстановлению, а также дифференцированию, переделке и пр. (М. Е. Лобашёв, 1967). Весьма доказательными были опыты Н. Тин-бергена (1969). Самки роющих ос обу чались распознавать круг из сосновых шишек, выложенных вокруг входа в норку (рис. 10). После отлета осы кольцо из шишек вокруг норки было перемещено в сторону. Возвратившись, оса ищет несуществующую норку внутри кольца и находит ее лишь после повторных облетов.
По-видимому, первые исследования научения у пчел проводились К. Фришем (1980), который показал, что пчелы при обучении запоминают запах цветка
| Таблица 3. Соотношение морфологических и функциональных ароморфозов на примере эволюции поведения беспозвоночных 1 (П.А.Тушмалова, 1987, с дополнениями) | ||
| Группы животных | Ароморфозы | |
| морфологический уровень (организация нервной системы) | функциональный уровень (высшая форма физиологических механизмов поведения) | |
| Простейшие | Донервный уровень | Привыкание, сснсит.изация |
| Кишечнополостные | Примитивная диффузная нервная система | Привыкание + тренированность |
| Плоские черви | Зачатки цефализации | Нестойкие условные рефлексы |
| Кольчатые черви | Ассоциативный центр мозга - грибовидные тела | Истинные условные рефлексы |
| Насекомые | Структурно-функциональная диф-ференцировка головного мозга | Положительные и отрицательные условные рефлексы, инструментальные условные рефлексы, цепные и комплексные условные рефлексы, условные рефлексы на отношения |
после первого прилета, окраску после 3—5 прилетов, а форму — примерно после 20 прилетов. Условно-рефлекторная же деятельность пчел в эксперименте изучалась М. Е. Лобашевым, которому удалось описать такие интегрирующие свойства мозга, которые ранее были обнаружены лишь у приматов (см. гл. VII). Хотя и медленнее, но и у пчел удалось выработать услов-^ ные рефлексы на трехчленные цепи 'зрительных стимулов, которые они начинали воспринимать как единый сигнал после 67— 158 сочетаний. У пчел обнаружены все виды внутреннего торможения, которое обеспечивает им высокий уровень аналитико-синтетической деятельности и достаточную пластичность поведения. У пчел допускают существование способности к элементарному обобщению и элементарному абстрагированию, что среди позвоночных присуще только высокоорганизованным млекопитающим.
Ф Таким образом, среди беспозвоночных формируется несколько форм обучения: привыкание, сенситизация, элементарные и истинные условные рефлексы, возникновение которых связано со значительным усложнением центральной нервной системы, они представляют принципиальное эволюционное приобретение — ароморфозы в общей динамике филогенетического развития механизмов поведения {табл. 3).
Позвоночные
Прежде чем перейти к анализу путей развития форм обучения у позвоночных, вкратце остановимся на рассмотрении особенностей поведения у самых низших хордовых — бесчерепных и круглоротых.
Бесчерепные.Б. Ф. Сергеевым (1967) показано, что у представителя бесчерепных — ланцетника центральная нервная система построена по типу нервной трубки, характеризуется экви-потенциальностью. т. е. отсутствием выраженной дифференциации и цефа-лизации, причем специализированные рецепторы еще не развиты. Если производить сочетание слабого условного раздражителя (света) с тем же светом, но большей интенсивности или на слабое тактильное раздражение, подкрепляемое электрическим ударом, то у этих животных удается выработать активную избегательную реакцию. Она сохраняется в течение 10 ч, угасая уже на вторые сутки, ее каждый день приходится вырабатывать заново. Создается впечатление о том, что у бесчерепных намечаются признаки ассоциативного условно-рефлекторного обучения, однако оно протекает по типу повышения возбудимости к условному раздражителю, т. е. сходно с сумма-ционными рефлексами.
У типичного представителя кругло-
poтых — речных миног впервые в эво-j люции формируются основные рецеп-торные системы и в зачаточном состоянии закладываются все подразделения мозга. Головной мозг состоит из конечного, промежуточного, среднего и ромбовидного вместе с мозжечком. В результате исследований А. В. Бару, В. В. Фанарджяна и Б. Ф. Сергеева было выявлено, что у миног вырабатываются оборонительные условные рефлексы после 3—4 сочетаний разномо-дальных сигналов (свет+электрический ток), эти реакции сохраняются в течение лишь одного дня, восстановление происходит самостоятельно или после нескольких подкреплений. Иными словами, хотя у круглоротых ассоциативное обучение носит характер условно-рефлекторной связи, этот тип реакций по ряду признаков напоминает еще суммационные рефлексы.
У пластинчатожаберных рыб дифференциация структур переднего мозга в морфофункциональном отношении еще недостаточно развита (А. И. Кара-мян, И. В. Малюкова, 1987). Это отражается и на характере условно-рефлекторной деятельности. У акул и скатов условные рефлексы вырабатываются медленно, осуществляются с большим латентным периодом, характеризуются непрочностью, значительной вариабельностью выполнения. Функцию высшего интегративного мозгового аппарата у этих животных выполняет сильно развитый мозжечок с его богатыми межцентральными связями с остальными отделами мозга. Передний мозг в условно-рефлекторной деятельности участия еще не принимает.
Костистые рыбы отличаются более сложной и дифференцированной организацией головного мозга и сенсорных систем. У них вырабатываются истинные пищевые и оборонительные условные рефлексы на простые и сложные световые, звуковые, обонятельные сигналы, постоянное и переменное магнитное поле. Костистые рыбы характеризуются всем репертуаром форм ассоциативного научения и сенсорного различения. Однако, как подчеркивает Б. Ф. Сергеев (1967), разобщенность
центральных аппаратов сенсорных систем, а также слабость нервных процессов'препятствует образованию внутри- и межсенсорных условных рефлексов (ассоциативные временные связи), что свидетельствует о низком уровне аналитико-синтетической деятельности.
В эволюции позвоночных рептилии занимают особое положение, ибо они представляют собой тот узловой этап эволюции, от которого формируются две независимые филогенетические линии развития мозга: стриарная, представленная птицами, и кортикальная, к которой относятся все млекопитающие вплоть до приматов и человека.
Мозг рептилий характеризуется прогрессивным усложнением структурной организации и впервые возникает примитивная таламокортикальная система связей (М. Г,. Белехова, 1977). Обнаружена слабая способность к фиксации следов выработанных условных рефлексов: при обучении новой задаче стирались следы предыдущего обучения. Стриатуму у черепах приписывается более важная роль в интегра-тивной деятельности мозга, чем дар-сальной коре, имеющей еще достаточно примитивную организацию.
Млекопитающие
Обратимся к филогенетическому древу (рис. 11), из которого видно, что по мере развития из рептилепо-добных предков основного ствола млекопитающих от него отходит ветвь современных однопроходных, позже сумчатых. Стволом же плацентарных млекопитающих являются насекомоядные, от которых произошли приматы, хищные и грызуны. Большая часть отрядов современных млекопитающих — результат параллельной эволюции, а не последовательной, к которой можно было бы отнести однопроходных, сумчатых, насекомоядных. В этом ряду и далее — вплоть до высших приматов и гоминид могут быть прослежены эволюционные преобразования мозга и поведения.
Филогенетический подход не исключает и другого, чрезвычайно важного

Рис. 11 Эволюционное дерево млекопитающих с выделением этапов филогенетической эволюции и уровней адаптивной эволюции (по А.С. Батуеву, 1979)
адаптационного подхода, который основан на изучении ныне живущих форм, обладающих разной степенью адаптации (специализации) в смысле развития мозга и сенсорных систем. Такие животные не обязательно должны иметь общую линию эволюции, и, следовательно, выводы, сделанные в результате такого сравнения, будут относиться только к общим принципам адаптации и выживания. Эти исследования углубляют понимание связи между развитием структурных образований мозга и их участием в целостном поведении. Филогенетический и адаптационный подходы взаимно дополняют друг друга. Поэтому правомерно говорить о филогенетической и адаптивной эволюции. Понятно, что сравнение особенностей поведения и конструкции мозга у грызунов, хищных, приматов не имеет никакого отношения к филогенетической эволюции, а лишь к адаптивной в смысле усложнения организации мозга *и усовершенствования биологических адаптации.
Начиная от насекомоядных можно проследить начальные этапы формиро вания всех таламических ядерных образований и проекционных зон нео-кортекса, характерных для высших млекопитающих. У насекомоядных (ежей) впервые в филогенезе формируется ассоциативная таламокортикальная система, которой присущи черты примитивной организации (А. С. Батуев, 1981) (см. гл. 7). Б. Ф. Сергееву (1967) удалось выработать у ежей ассоциативные временные связи возбудительного и тормозного характера путем сочетания разномодальных звуковых и зрительных раздражений. И. В. Милюкова (1981) установила у ежей с помощью сложных .двигательных условных рефлексов различение освещенности, полос разной ориентации, фигур. Такие условные рефлексы вырабатывались быстрее, чем у черепах, и сохранялись в течение 40—50 дней. Научившись новой задаче, ежи не забывали предыдущих навыков, что свидетельствует о большой прочности хранения следов.
Однако условные рефлексы на одновременный и последовательный комплексные раздражители, которые вырабатывались с трудом, часто сопровождались невротическими состояниями. В отличие от высших млекопитающих у ежей слабо развиты ориентировочно-исследовательские реакции, не наблюдается использования метода «проб и ошибок», их нервные процессы характеризуются инертностью. Наряду с участием примитивной ассоциативной коры в процессах сенсомоторного синтеза основная интегративная роль в об-щсмозговом контроле за сложными формами поведения принадлежит подкорковым отделам переднего мозга— стриарным образованиям. По этому признаку насекомоядные еще сохраняют черты, присущие рептилиям.
Приматы
Важнейшим показателем эволюционного прогресса нервной системы является развитие неокортекса, а в нем— ассоциативных кортикальных полей и соответствующих ядер таламуса (А. С. Батуев, 1981). Именно эти структуры мозга достигают у обезьян высокого уровня развития. В самой коре впервые в эволюции формируются длинные пучки волокон, связывающие различные области коры между собой. Все это создает предпосылки для выполнения весьма сложных форм поведения.
Значительное место в жизни обезьян имеет ориентировочно-исследовательская деятельность, которая, по мысли Я. Дембовского (1965), является зачатком будущего интеллекта. Убедительно показано, что обезьяны способны к формированию самых сложных форм условно-рефлекторной деятельности, связанной с тонким различением зрительных объектов по самым разнообразным признакам. Исследования следовых условных рефлексов выявили прогресс образной памяти. Длительность хранения следов у низших обезьян исчисляется месяцами н годами. Обезьянам присущи все формы когнитивного обучения: образное поведение, рассудочная деятельность и вероятностное прогнозирование.
Все вышесказанное позволило А. И. Карамнн\ (1976), А. И. Карамяну и И. В. Малкжовой (1987) выдвинуть представление о существовании критических этапов развития мозгового обеспечения поведения в эволюции позвоночных.
Первый критический этап представлен бесчерепными и круглоротыми, характеризующимися еще слабой дифференциацией головного мозга (рис. 12). Так, у ланцетников (бесчерепные) удается выработать лишь суммаиионные реакции, а у миног (круглоротые)— квазиусловные рефлексы, занимающие промежуточное положение между сум-мационными и истинными рефлексами.
Второй критический этап представлен пластиножаберными рыбами (скаты, акулы), структуры таламуса и конечного мозга которых еще недостаточно развиты. У этих животных удалось выработать условные рефлексы, которые были непрочными, сохранялись в течение нескольких суток и характеризовались невысоким уровнем их осуществления. Передний мозг пластино-жаберных рыб не принимает участия в организации сложных поведенческих актов.
Третий этап представлен рептилиями (особенно черепахами), на поверхности конечного мозга которых формируется примитивный неокортекс. Эти животные способны к формированию более сложных форм поведения по сравнению с низшими позвоночными. Однако высшая нервная деятельность черепах отличается слабостью нервных процессов, несовершенством памяти, отсутствием способности к вероятностному прогнозированию. Ведущую роль в интегративной функции мозга у черепах выполняет полосатое тело (стриатум).
Четвертый критический этап представлен низшими млекопитающими — насекомоядными. Мозг ежей характеризуется увеличением общей поверхности неокортекса и развитием дорзаль-ных ядер таламуса. Однако четкая дифференциация структур таламуса и коры выражена еще слабо, хотя обна-
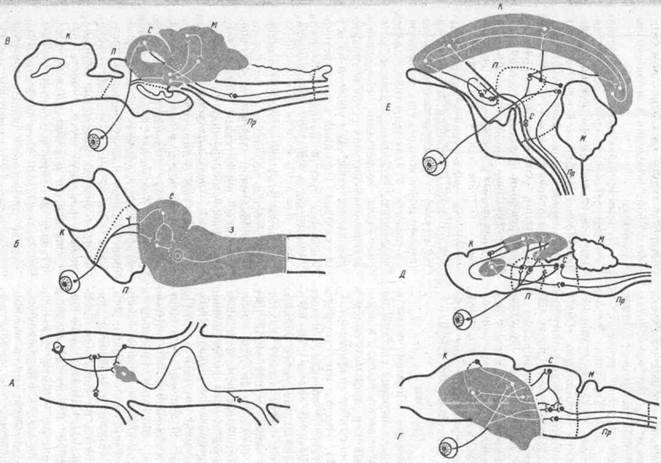
Рис. 12. Этапы развитияуровней интеграции в филогенезепозвоночных ( по А. И. Карамяну, 1976):
а — бесчерепные (ланцетник), спинальный уровень интеграции, в центре гигантская клетка с аксоном, обеспечивающим нисходящие, связи; б — круглоротые (минога), бульбомезэнцефалнческий уровень интеграции; в — пластинчатожаберные рыбы (акула), мезэнцефалоцеребел-ляриый уровень интеграции; г — рептилии (черепаха), диэнцефалютелэнцефальный уровень интеграции; д — насекомоядные (еж), стр но кортикальный уровень интеграции; е — приматы (обезьяна), неокортикальнын уровень интеграции; заштрихованы высшие интегративные уровни мозга; К — конечный мозг; П — промежуточный; С — средний; 3 — задний; /И — мозжечок; Пр — продолговатый мозг. На схеме показаны только зрительные афферентные проекции. Относительные размеры мозга разных животных не соблюдены
ружено существование примитивной ассоциативной системы.
У ежей образуются сложные последовательные цепи двигательных поведенческих актов при зрительном распознавании. Впервые в эволюции позвоночных проявляется хотя и в слабой форме свойство вероятностного прогнозирования. Высшая нервная деятельность ежей характеризуется примитивностью: инертностью нервных процессов, трудностью формирования межсенсорных синтезов, слабостью ориентировочно-исследовательских реакций.
Пятый критический этап представлен приматами, главной особенностью организации мозга которых является мощное развитие его ассоциативных систем, объединенных кортикокорти-кальными путями в единую интегра-тивную систему мозга. Таким образом создаются все предпосылки для формирования более сложных форм поведения, которые оказываются специфически присущими лишь приматам.
Высшая нервная деятельность человекообразных обезьян.Некоторые исследователи поведения животных считают, что высшие психические функции не что иное, как примитивное мышление, которым животные могут пользоваться без предварительного научения, т. е. не используя прежний жизненный опыт. Так, В, Келлер после наблюдения за поведением шимпанзе полагал, что высшие психические функции являются выражением изначальных свойств и способностей мозга. И, П. Павлов и его сотрудники, повторив опыты Келлера, пришли к иному выводу. Шимпанзе составляет пирамиду из ящиков, чтобы достать гроздь бананов, подвешенных к потолку, на основании .перенесения в новую ситуацию старого опыта, на основании проб и ошибок (рис. 13). Сам И. П. Павлов писал по этому поводу; «...Каждая новая ассоциация, касающаяся отношения внешних вещей, есть прибавление знания, а пользование этим знанием есть то, что называется пониманием»1.
'Павлов И. П. Неопубликованные и малоизвестные материалы И. П. Павлова. Л., Наука, 1975. С. 96.
Еще Ч. Дарвин (1871) отмечал, что подражание сыграло значительную роль в процессе антропогенеза, причем у родоначальников человека подражание в сочетании с рассудком сыграли исключительную роль в выборе оптимальных решений.
Л. А. Фирсов (1987) выделяет подражательную деятельность в качестве самостоятельной формы поведения антропоидов, которая оказывается особенно выраженной у денетышей шимпанзе в возрасте 2—2,5 лет. Причем подражание после однократного наблюдения решения проблемных задач или демонстрации дифференцирования предъявляемых сигналов особью-«ак-тером» сохраняется у «зрителя» от нескольких часов до нескольких недель. Причем кроме скорости и прочности подражания отмечается его очень широкий диапазон.
Механизм подражания следует рассматривать как особую форму поведения или отражения действительности, где принцип «животное —- окружающий мир» заменяется принципом «животное — другое животное — окружающий мир». Причем особь-«актер» воспринимается особью-«зрителем» как обобщенная система натуральных раздражений, которыми и являются акты поведения.
Предметная и орудийная деятельность антропоидов — это одна из базовых проблем антропогенеза. Установлено, что манипулирование разнообразными предметами среды — предметная деятельность — это неотъемлемая черта сложного поведения всех представителей отряда приматов. Антропоиды обладают способностью к целенаправленному манипулированию с предметами. В процессе этой деятельности они не только используют предмет как таковой или при необходимости изменяют его, но способны также сочленить несколько предметов в устойчивую конструкцию. Использование предметов для игры молодыми шимпанзе начинается с 2,5—3-месячного возраста. Но только в возрасте около 2,5 лет детеныши шимпанзе научались составлять новые конструкции и использо-

Рис 13 Решение шимпаезе задачи получения банана (по В. Детьери Э. Стеллар, 1967)
вать их для приближения приманки. То же наблюдается и у других антропоидов (рис. 14).
Орудийная деятельность отчетливо проявляется у шимпанзе в опытах с огнем, который служил препятствием для достижения приманки. В опытах И. П. Павлова Рафаэль взял бутылку с водой и вылил ее на пламя, а в опытах Л. А. Фирсова Каролина накрыла огонь половой тряпкой и легко завладела апельсином. Из этих опытов ясно
видно свойство обобщения сигнальной ценности окружающих предметов. Наблюдения за поведением шимпанзе, проведенные Л. А. Фирсовым, дают основание считать, что, прежде чем начать какие-либо действия, у шимпанзе имеется ближайший план, ради которого они реализуются. Вот один из примеров. Подростки шимпанзе Лада и Нева, воспользовавшись ошибкой лаборантки, достали связку ключей, забытую ею на столе в 3 м от вольера, и оказались на свободе. Действия обезьян при этом были следующими: 1) отгрызание края столешницы от стола, который уже давно находился в вольере; 2) с помощью получившейся палки подтягивание занавеси и овладение ею; 3) набрасывание как лассо занавеси на стол с ключами и очень осторожное подтягивание связки ключей к вольере; 4) открывание висячего замка и выход па свободу.
Этот пример отражает реализацию уже принятого окончательного решения, которое сформировалось как па.основе текущей деятельности, так и имеющегося у обезьян жизненного опыта. В любом случае обезьяна способна в известной степени предвидеть результаты своих действий.
Орудийная деятельность основана на анализе связей между предметами, явлениями и результатами манипулирования с предметами. При этом важную роль играет уровень обобщения элементов среды с тем, чтобы можно было применить любой предмет в любой ситуации, придерживаясь логики принятого решения. Целенаправленность поведения с обобщенным использованием предметов внешней среды, с частичным их изменением и составлением из них устойчивых конструкций является наиболее поздним эволюционным приобретением. Она обязательно проявляется в повелении ребенка и на ней в значительной мере основывается трудовая деятельность человека. Новая ступень предметной деятельности — орудийная деятельность — развивается в процессе все большей социализации древнейшего человека,
Учитывая семейно-групповой образ

Рис. 14. Орангутан за составлением двух трубчатых предметов (по Л. А. Фнрсову, 1987)
жизни антропоидов, исследователи уделяют значительное внимание способам общения животных между собой и с экспериментатором. Основное внимание уделяется голосовым и жести кулятор-ным проявлениям. Л: А. Фирсов и В. В. Плотников (1981) отмечают выраженную «полисемантичность» голосовых реакций обезьян и большое значение окружающей ситуации для придании им определенной сигнальной значимости. В силу высокоразвитых свойств подражания антропоиды легко усваивают новые звуковые проявления, обогащая их экологическое значение. В определенных условиях эти артифици-альныс сигналы могли оказаться более точными, чем врожденные, и соответственно их биологическая роль возрастала.
Своеобразная форма общения шимпанзе с экспериментатором была изучена А. И. Счастным (1972). Обезьян приучали возвращать экспериментатору выданные им во время опыта жетоны: за возвращение круглого жетона давали обезьяне конфету, квадратного — орех, прямоугольного — компот, шестиугольного — игрушку, за треугольник — ничего не давали. Обезьяны, используя условные «знаки», сигнали-
зировали экспериментатору и друг другу о своей потребности в пище, питье или игрушке.
Та же цель преследовалась опытом супругов Гарднеров, Премаков и других исследователей на детенышах шимпанзе и горилл. В качестве символов могли использоваться либо специальные фишки, либо азбука глухонемых. Важно то, что выученными жестами или фишками шимпанзе могли пользоваться между собой, а новички по механизму подражания воспринимали их у обученных и успешно ими пользовались.
Антропоиды обладают способностью к переносу первичного научения для формирования обобщения с отвлечением от конкретных признаков сигнала. Стало очевидным, что шимпанзе способны абстрагироваться от некоторых существенных признаков предметов или явлений и обнаруживают выраженную способность к ассоциативному мышлению. Обобщение и абстракция являются результатом чувственного опыта животного, содержат информацию об истории процесса, оценке ситуации, решении, его реализации, результатах комбинаторики старых навыков для нового решения. Поэтому обобщение — функ,-

циональный блок систематизированной информации, хранящейся в аппаратах памяти.
Вероятно, на основе сложных обобщений у древних приматов формировался знаковый язык. Л. А. Фирсов (1982) понимает под термином «язык» все формы адаптивного поведения животных и человека, в том числе врожденные коммуникативные сигналы. Это первичный язык. Знаковая система общения у обезьян позволяет различать во вторичных языках стадию «А» (довербальные понятия) и стадию «Б» (вербальные понятия). Последние, как правило, реализуются с помощью ре-
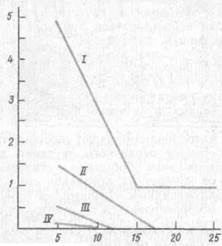
Рис. 15. Состояние образной памяти (отсроченный выбор) на фоне трениров-
ки различения зрительных сигналов (по Л. А. Фирсову, 1987):
по осиординат — отсрочка в критическом испытании, мин; по оси абсцисс —номера критического испытания после образованияусловного рефлекса; / — шимпанзе; //— павианы, /// —- макаки; IV — капуцины
чевого аппарата. Соотношение классификации языков приводится в табл. 4.
Существенной стороной поведения антропоидов являются формы их сосуществования в сообществах (семьях, группах, стадах). Такую кооперацию можно рассматривать как присущий приматам тип зоосоциального поведения, при котором действия нескольких особей объединяются для выполнения какой-либо операции, направленной на достижение полезного результата. Кооперативность поведения обезьян зависит прежде всего от иерархических рангов партнеров, а затем и от других факторов, таких, как уровень мотивации, физиологическое состояние особей и др.
Кроме обычного наблюдения за поведением животных Л. А. Фирсовым был использован весьма показательный объективный тест на кооперативное поведение двух обезьян. Возле одной из кормушек находился рычаг, нажатие на который открывало доступ к пище в противоположной кормушке, и наоборот. Кооперативное поведение устанавливается тогда, когда обезьяны по очереди подкрепляют одна другую. Однако такое поведение разрушалось, как только проявилось доминирование одной особи и подчиненность другой. В этом случае подчиненная особь в несколько раз чаще нажимала на рычаг, чтобы накормить своего партнера, чем доминирующая.
Наконец, хорошо изучена способность антропоидов к длительному хранению следов. Главной отличительной особенностью шимпанзе является широкое использование образной памяти (рис. 15).
Вышеприведенные данные об уровне психического развития антропоидов, включающего подражательную, орудийную, ориентировочно-исследовательскую деятельность, способность к обобщению и абстрагированию сигналов, использованию знаковой системы сигнализации и высокого уровня образной памяти, позволяют заключить, что обезьянообразные, ближайшие предшественники древнейшего человека, обладали не меньшим развитием перечисленных функций.
Дата добавления: 2015-08-04; просмотров: 1024;