ВОЗРАСТНАЯДИНАМИКА ГЕНОТИП-СРЕДОВЫХ СООТНОШЕНИЙ В ЭЛЕКТРОЭНЦЕФАЛОГРАММЕ
Несмотря на то что темпы созревания биоэлектрической активности, проявляющиеся в возрастании основной частоты и регулярности, высоко индивидуальны, более высокое сходство ЭЭГ в парах МЗ близнецов по сравнению с ДЗ характерно не только для взрослых, но и для детей. Ф. Фогель в результате изучения возрастной динамики внутрипарного сходства 110 пар МЗ и 96 пар ДЗ близнецов с 6 до 80 лет пришел к заключению, что темп возрастных изменений ЭЭГ обусловлен генетически, поскольку в периоды ее наиболее заметной перестройки — ив детстве, и в зрелом возрасте — МЗ близнецы имеют идентичные кривые [159]. Однако специального анализа возрастных различий в наследуемости параметров ЭЭГ он не проводил.
Исследований, прямо посвященных изучению возрастной динамики генотип-средовых соотношений в изменчивости ЭЭГ и ее параметров, немного. Они появились в начале 70-х годов и были выполнены методом близнецов [88, 162]. В них целенаправленно сравнивались эффекты генетических влияний в межиндивидуальной вариативности параметров ЭЭГ на разных этапах онтогенеза. Наиболее интересный результат заключался в том, что генетические влияния в параметрах ЭЭГ в разных возрастах проявлялись с разной силой. Иными словами, в одних случаях эти влияния были выражены сильнее, в других слабее, в третьих не обнаруживались совсем. Так, при сопоставлении данных близнецов трех возрастных групп (младших школьников, подростков и взрослых) были выявлены значительные возрастные различия в генотип-средовой детерминации параметров ЭЭГ
25* 387
Таблица 18.1
Внутриклассовые корреляции параметров ЭЭГ покояу близнецов
трех возрастов [по: 33]
| Возраст (годы) | Группа | Суммарная энергия | ||||
| дельта | тэта | альфа | бета-1 | бета-2 | ||
| 10-11 14-16 18-25 | МЗ (п = 30) ДЗ (п = 26) М3(n=19) Д3(n=19) МЗ(n=26) ДЗ(n=22) | 661**5 73** 735** 655** 584** 288 | 777** 537** 852** 660** 855*** 409 | 807*** 528** 949*** 426* 900*** 663** | 890*** 793** 862*** 267 759** 491* | 712** 180 633** 208 850*** 282 |
| Частота | ||||||
| 10-11 14-16 18-25 | МЗ (п = 30) ДЗ(n=26) МЗ(n=19) ДЗ(n=19) МЗ (п = 26) ДЗ(n=22) | 651** 066 114 -131 -140 -024 | 848** * о 1 z^sksk 180 -098 520** 019 | 873*** 483* 754** 622* 433* | 242 379 405 -090 | 501* 506* 653* 318 |
* р < 0,05; **р < 0,01; *** р < 0,001.
(табл. 18.1). По этим данным можно, например, констатировать некоторое увеличение наследуемости суммарной энергии альфа- и бета-1 ритмов в подростковом возрасте. Однако относительно небольшое количество пар не позволяет считать этот факт окончательно установленным.
В исследовании 25 пар МЗ и 12 пар ДЗ близнецов в возрасте 10-12 мес. анализировался вклад генотипа в индивидуальные особенности доминирующей частоты 6-8 Гц, которая рассматривалась авторами как функциональный аналог альфа-ритма [499]. Показатель наследуемости составил 0,67. В 10-11 лет показатель наследуемости частоты альфа-ритма был 0,78, а во взрослом возрасте — 0,60 [132], Таким образом, вклад генотипа в индивидуальные особенности частоты альфа-ритма относительно высок на разных этапах развития. Тем не менее сравнение показателей наследуемости не позволяет установить, идентичные или разные генетические влияния определяют межиндивидуальную изменчивость данного показателя в изученные периоды развития. Для установления генетически обусловленной преемственности
развития необходимо лонгитюдное исследование наследуемости параметров альфа-ритма,
Данный подход получил продолжение в последние годы в исследованиях, выполненных на значительно больших выборках близнецов и с привлечением современных методов генетико-статистического анализа. Наиболее представительной в этом плане является программа исследований, проводимых голландскими учеными.
В исследовании К. Ван Баал [430] на материале 209 пар МЗ и ДЗ близнецов 5 лет изучались генотип-средовые соотношения в спектральной мощности ЭЭГ по шести частотным диапазонам (табл. 18.2). Оценивали абсолютную суммарную мощность в соответствующих диапазонах и относительную мощность, определяемую как отношение мощности каждого диапазона к общей суммарной мощности. Относительная мощность характеризует степень выраженности данной спектральной составляющей в общем спектре ЭЭГ.
Таблица 18.2
Наследуемость спектральных мощностей ЭЭГ покоя в пятилетнем возрасте [по: 430]
| Спектральная полоса | Частотный диапазон (в Гц) | Наследуемость (в %) (усредненная по 14 отведениям) | |
| абсолютная мощность | относительная мощность | ||
| дельта тэта альфа-1 альфа-2 бета-1 бета-2 | 1,5-3,5 4,0-7,5 8,0-9,5 10,0-12,5 13,0-17,5 18,0-25,0 | 81 81 78 78 64 | 76 71 72 68 65 |
В результате было установлено более высокое сходство МЗ близнецов по всем ритмическим составляющим спектра ЭЭГ (его абсолютной и относительной мощности) в полосах: дельта, тэта, альфа-1, альфа-2, бета-1, бета-2. Генетическая модель, включающая аддитивный генетический и уникальный (индивидуальный) средовый компоненты, объясняет индивидуальные различия как в абсолютной, так и в относительной мощности почти во всех спектральных полосах и во всех точках регистрации ЭЭГ. Оценка влияний систематической среды не проводилась. Наследуемость спектральных составляющих ЭЭГ по показателям большинства полос оказалась очень
высокой (см. табл. 18.2). Влияния факторов пола обнаружено не было, Эти факты говорят о том, что в 5 лет индивидуальные особенности фоновой ЭЭГ в значительной степени обусловлены генотипом.
Несколько иные результаты были получены при исследовании генетической обусловленности трех субкомпонентов альфа по показателям спектральной мощности на близнецах 6-8 лет [32]. Три поддиапазона включали: альфа-1 - 7,7-8,9 Гц; альфа-2 - 9,3-10,5 Гц; альфа-3 - 10,9-12,5 Гц [143]. Исследование генетической обусловленности трех субкомпонентов альфа-, проведенное на близнецах 6-8 лет методом подбора моделей, показало, что в этом возрасте индивидуальные различия спектральных характеристик низкочастотной составляющей обусловлены в основном аддитивными генетическими эффектами, а высокочастотной составляющей — эффектами систематической и случайной среды. Таким образом, функциональная неоднородность альфа-ритма в этом возрасте, видимо, имеет определенные генетические основания.
В рамках упоминавшейся выше голландской программы, т.е. с использованием тех же методов регистрации и способов анализа, проводилось изучение наследуемости спектральных характеристик ЭЭГ подростков 16 лет (91 пара МЗ и 122 пары ДЗ близнецов), Практически по всем показателям наблюдалось высокое сходство МЗ близнецов; коэффициенты внутриклассовой корреляции у них составили около 0,85, у ДЗ — в большинстве случаев в два раза меньше; коэффициенты наследуемости у подростков в среднем несколько выше, чем у 5-летних детей. Так, усредненный по всем отведениям коэффициент наследуемости составляет 0,90, лишь в дельта-диапазоне он несколько ниже — 0,75. Существенных межзональных различий не установлено. Как и в исследовании 5-летних близнецов, наиболее соответствующей оказалась простая генетическая модель [432].
К. Ван Баал с коллегами [430] изучали также наследственные влияния в когерентности ЭЭГ, которая расценивается как показатель интенсивности связей, существующих между разными отделами мозга. Данные были получены при исследовании фоновой ЭЭГ у 167 пар МЗ и ДЗ близнецов 5 лет. Когерентность вычислялась в тэта-полосе 4,0-7,5 Гц, поскольку тэта-ритм в этом возрасте считается доминирующим ритмом ЭЭГ. Когерентность оценивалась между префронталь-ной, фронтальной, центральной, теменной и затылочной областями отдельно в каждом полушарии (рис. 18.1), причем в двух вариантах — для коротких и длинных расстояний. В первом случае оценка когерентности проводилась попарно для тэта-ритма префронтальной и фронтальной, пре фронтальной и центральной, центральной и затылочной, центральной и теменной, теменной и затылочной областей (расстояние между точками отведения 7-14 см). Во втором случае попарно сравнивался тэта-ритм фронтальной и затылочной зон, префронталь-ной и теменной, префронтальной и затылочной (расстояние между точками отведения 21-28 см).
Рнс 20.1.Наследуемость когерентности (аддитивная и доминантная составляющие с 30% доверительным интервалом).
а—для префронтальныхотведений^.и/1,; б — для затылочных отведений Ли О,
[по: 430J.
Обозначения: □ — аддитивная наследуемость; ■— доминантная наследуемость.
Генетический анализ, проведенный методом подбора моделей, обнаружил значительные влияния генетических факторов во всех показателях когерентности. Наследуемость в широком смысле оказалась довольно высокой в разных вариантах сопоставления отведений. Средняя величина наследуемости составляла 49%, варьируя в диапазоне от 30 до 71%. При этом был установлен ряд интересных фактов, и прежде всего — различия в наследуемости для показателей когерентности, полученных на коротких и длинных расстояниях. Когерентность тэта-ритма для соседних отведений (например, префронтального и фронтального, теменного и затылочного) обнаружила значительно меньшую наследуемость по сравнению с когерентностью, характеризующей взаимосвязь далеко расположенных зон (префронтальной и затылочной, префронтальной и теменной). Этот факт представляется особенно существенным, если иметь в виду, что ретестовая надежность выше на коротких расстояниях: ее показатель варьирует от 0,91 для соседних отведений до 0,62 для наибольшего расстояния.
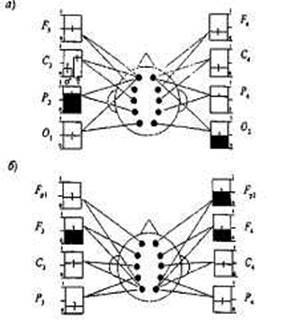
Таким образом, хотя ошибка измерения возрастала по мере увеличения расстояния между отведениями, показатель наследуемости не только не становился ниже, а, напротив, увеличивался. Невысокая наследуемость когерентности в соседних отведениях в сочетании с высокой надежностью говорит о существенном вкладе среды в формирование когерентных отношений между тэта-активностью соседних зон. Что касается когерентности тэта-ритма на длинных расстояниях, есть все основания говорить о значительном вкладе генетических факторов в межиндивидуальную изменчивость по этим показателям. Причем генетический компонент здесь в основном формируется за счет доминантной составляющей, в то время как наследуемость когерентности близких отведений полностью определяется только аддитивными влияниями (см. рис. 18.1).
По-видимому, длинные аксональные связи между сенсорными проекционными и фронтальными областями в своем формировании больше зависят от генотипа, чем межнейронные связи в соседних зонах мозга.
В дальнейшем онтогенезе происходят существенные изменения в спектре ЭЭГ и когерентности. Закономерно возникает вопрос: как проявляют себя генетические влияния в этих показателях на следующих стадиях развития? Для того чтобы оценить природу межиндивидуальной изменчивости возрастных преобразований когерентности, те же авторы исследовали когерентность тэта-ритма у тех же близнецов по прошествии в среднем 1 года 7 месяцев после первой регистрации. 192 пары близнецов прошли повторное исследование в 7 лет. Общая схема регистрации ЭЭГ и подсчета когерентности в обеих возрастных группах совпадали, что обеспечивало возможность такого анализа.
Авторы ставили две задачи: во-первых, установить, увеличиваются ли с 5 до 7 лет генетические влияния в индивидуальных особенностях когерентности ЭЭГ, т.е. будет ли наследуемость по этим показателям в 7 лет выше, чем в 5, и, во-вторых, выяснить, включаются ли новые генетические влияния в 7 лет, или генетические эффекты опосредуются одними и теми же генами и в том, и в другом возрасте. Методом подбора моделей в межвозрастной вариативности и ковариа-ции когерентности в 5 и 7 лет были выделены средовая и генетическая составляющие, общие для обоих возрастов и специфические для 7 лет. Динамика показателей наследуемости, генетического и средового (уникального для каждого члена пары) компонентов дисперсии представлена в табл. 18.3. Оказалось, что в 7 лет наследуемость когерентности в передних отделах коры снизилась, а в задних, напротив, возросла (в том и другом случае в среднем на 10-15%). Средовый компонент дисперсии (влияния индивидуальной среды) с возрастом уменьшился.
Тенденция к увеличению наследуемости когерентности с возрастом подтверждается данными и другого исследования, в котором изучалась внутриполушарная когерентность у 213 пар близнецов подрост-
Таблица 18.3
Дата добавления: 2015-07-18; просмотров: 967;