ОРГАНЫ СВЕЧЕНИЯ
В кожном покрове рыб различают два слоя: наружный слой эпителиальных клеток, или эпидермис, и внутренний слой из соединительно тканных клеток – собственно кожа, дерма, кориум, кутис (рис. 5).
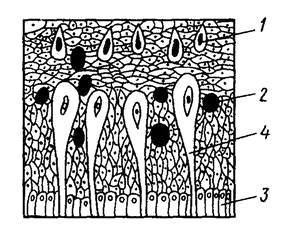
Рис. 5. Строение кожи у рыб (усач) (по Строганову, 1962):
1 – бокаловидные клетки; 2 – зернистые, 3 – базальные, 4 – колбовидные
Кожа подстилается рыхлой соединительнотканной прослойкой (подкожная соединительная ткань, подкожная клетчатка). У многих рыб в подкожной клетчатке откладывается жир.
Эпидермис защищает организм от проникновения в него веществ извне. Он состоит из многослойного эпителия, форма клеток и количество слоев которого варьируют у разных рыб. Самый наружный слой эпителиальных клеток ороговевает, но в отличие от наземных позвоночных у рыб он не отмирает, сохраняя связь с живыми клетками. В течение жизни рыбы интенсивность ороговения эпидермиса не остается неизменной, наибольшей степени оно достигает у некоторых рыб перед нерестом: так, у самцов карповых и сиговых в некоторых местах тела (особенно на голове, жаберных крышках, боках и т. д. ) появляется так называемая жемчужная сыпь – масса мелких белых бугорков, придающих коже шероховатость. После нереста она исчезает.
Кожа рыб отличается от кожи других позвоночных большим количеством слизи.
Слизь образуется в специализированных железистых клетках, лежащих в эпидермисе. Это клетки трех форм: бокаловидные, колбовидные и зернистые. Присутствие всех форм определяет наибольшее количество слизи, при наличии одного вида клеток слизи выделяется значительно меньше.
Интенсивность выделения слизи у разных рыб различна. Как правило, рыбы с хорошо развитым чешуйчатым покровом выделяют слизи меньше (лососевые, окуневые).
Рыбы, лишенные чешуи или чешуя которых редуцирована (круглоротые, некоторые сомы, линь, вьюн), выделяют слизи очень много. С бактерицидными свойствами слизи линя связывают повышенную устойчивость его к заражению паразитами, тогда как другие представители семейства карповых такой устойчивостью не обладают.
Роль слизи в жизненных отправлениях рыб очень велика: она уменьшает трение тела о воду (механическая защита), предотвращает попадание в организм паразитов и бактерий (бактерицидная защита), ускоряет свертывание крови в случаях ранений, способствует выведению веществ из организма, регулирует проникновение воды и солей (осмотическая регуляция), осаждает муть и выделяет специфический видовой запах.
По некоторым данным, слизь некоторых рыб ядовита. Слизь миног вызывает у хищников нарушение пищеварения.
Также в одноклеточных железах кожи образуются феромоны –летучие (пахучие) вещества, выделяемые в окружающую среду и воздействующие на рецепторы других рыб. Они специфичны для разных видов, даже близкородственных; в некоторых случаях определена их внутривидовая дифференцировка (возрастная, половая).
У многих рыб, в том числе у карповых, образуется так называемое вещество страха (ихтиоптерин), которое выделяется в воду из тела пораненной особи и воспринимается ее сородичами как сигнал, извещающий об опасности.
Светящиеся органы морских глубоководных рыб и ядовитые железы некоторых рыб, помещающиеся у основания шипов и плавниковых лучей (морской дракон), также являются железистыми образованиями эпидермиса.
В нижних слоях эпидермиса в пограничных с ними слоях кориума залегают пигментные клетки– хроматофоры. Хроматофоры –звездчатые клетки, со множеством отростков, включающие зернышки пигмента. Они определяют все разнообразие окраски рыб, особенно яркой в тропиках. Различие в окраске достигается сочетанием разных хроматофоров: меланофоры имеют зерна черного пигмента, ксантофоры – жёлтого, эритрофоры – красного; гуанофоры или иридоциты не имеют пигментных зерен, но содержат кристаллики гуанина, благодаря которым рыба приобретает серебристую окраску.
Интенсивность окраски определяется состоянием хроматофоров: при их расширении пигментные зерна растекаются на большее пространство и окраска тела становится яркой. Если хроматофоры сокращаются, пигментные зерна скапливаются в центре, оставляя большую часть клетки неокрашенной (рис. 6), и окраска тела бледнеет.
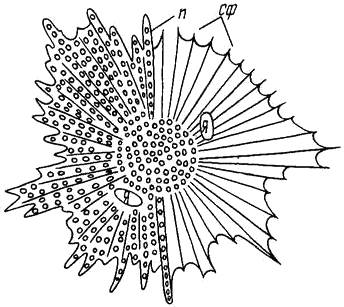
Рис. 6. Меланофор карася.
Справа – фаза контракции, слева – фаза экспансии (по Строганову, 1962):
я – ядра, п – пигментные зёрна, сф – скелетная фибрилла
Форма пигментных клеток постоянна и у близких видов сходна.
Состояние пигментных клеток меняется в зависимости от внешних и внутренних факторов: температуры и газового режима водоема, возраста, пола, состояния организма (голод, размножение и т. п. ), эмоций (возбуждение, страх) и т. д.
Окраска тела часто имеет приспособительное значение. У пелагических рыб обычно спина темная, а брюшко серебристо-белое, из-за чего рыба малозаметна и сверху и снизу.
Общеизвестно приспособление окраски тела к цвету дна у камбал, бычков, морских игл: они могут повторять даже рисунок шахматной доски, положенной на дно. При этом основную роль играет нервная система, воспроизводящая зрительные раздражения и вызывающая соответствующую реакцию пигментных клеток (у ослепленных рыб кожа не меняла окраску).
Изменение окраски в период размножения (появление брачного наряда), часто наблюдаемое в этот период различие в окраске самок и самцов происходят под влиянием гормональных факторов.
Собственно кожа обеспечивает прочность покровов. Она состоит из нескольких слоев соединительной ткани (у костистых рыб, например, из трех), пронизанных нервами и капиллярами. В этом же слое залегают специализированные клетки – склеробласты, выделяющие секрет, который, застывая, образует чешую (кожный скелет), основным назначением которой является механическая защита тела.
Все настоящие рыбы обладают чешуей, но степень развития её у разных рыб различна. Некоторые рыбы (например, сомы, скаты)в процессе эволюции утратили чешую, так что отсутствие ее у них является вторичным явлением.
Размер чешуек сильно варьирует – от микроскопических у угрей до нескольких сантиметров у индийского усача. Также разнообразна форма чешуек.
Различают три формы чешуи (рис. 7): плакоидная, ганоидная и костная. Плакоидная – наиболее древняя, сохранилась у хрящевых рыб (акулы, скаты). Состоит из пластинки, на которой возвышается шипик. Старые чешуйки сбрасываются, на их месте возникают новые. Ганоидная – преимущественно у ископаемых рыб. Чешуйки имеют ромбическую форму, тесно сочленяются одна с другой, так что тело оказывается заключенным в панцирь. Чешуйки со временем не меняются. Названием своим чешуя обязана ганоину (дентинообразному веществу), толстым слоем лежащему на костной пластинке. Среди современных рыб ее имеют панцирные щуки и многоперы. Кроме того, она имеется у осетровых в виде пластинок на верхней лопасти хвостового плавника (фулькры) и жучек, разбросанных по телу (модификация нескольких слившихся ганоидных чешуек). Постепенно видоизменяясь, чешуя теряла ганоин. У современных костистых рыб ганоина уже нет, чешуйки состоят из костных пластинок (костная чешуя). Эти чешуйки могут быть: циклоидными –округлыми, с гладкими краями (карповые) и ктеноидными – с зазубренным задним краем (окуневые). Обе формы родственны, но циклоидная как более примитивная встречается у низкоорганизованных рыб. Бывают случаи, когда в пределах одного вида самцы имеют ктеноидную, а самки – циклоидную чешую (камбалы рода Liopsetta), или даже у одной особи встречаются чешуйки обеих форм.
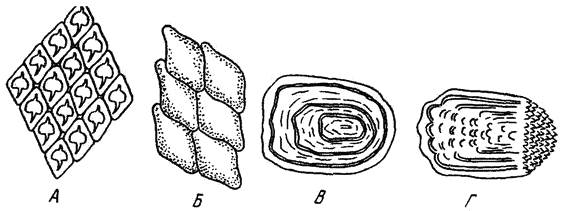
Рис. 7. Форма чешуи рыб.
А – плакоидная; Б – ганоидная; В – циклоидная; Г – ктеноидная
Каждая такая чешуйка начинает образовываться в чешуйном кармашке – углублении в кориуме, затем свободный конец ее выходит из кармашка и накладывается на следующую чешуйку. Такое черепицеобразное расположение чешуек позволяет телу рыбы свободно изгибаться. Чешуя располагается рядами. Число рядов и количество чешуи в продольном ряду не изменяются с возрастом рыбы, поэтому они служат показателями при систематическом определении.
В разрезе на каждой чешуйке различаются наружный покрышечный слой (преимущественно неорганического состава) и нижний базальный (имеющий характер кости). На верху покрышечного слоя концентрическими слоями располагаются ребрышки – склериты. Образование склеритов происходит периодически, поэтому их ряды подсчитывают при определении возраста рыб (годичные кольца) и обратного расчисления роста (см. с. 117).
Кожа рыб быстро регенерирует. Через кожу происходит, с одной стороны, частичное выделение конечных продуктов обмена веществ, а с другой – поглощение некоторых веществ из внешней среды (кислород, угольная кислота, вода, сера, фосфор, кальций и другие элементы, играющие большую роль в жизнедеятельности). Большую роль играет кожа как рецепторная поверхность: в ней располагаются термо-, баро- хемо- и другие рецепторы. В толще кориума образуются покровные кости черепа и пояса грудных плавников.
Скелет
Скелет у большей части рыб двоякий: наружный – защитный (чешуя) и внутренний – опорный.
Внутренний скелет состоит из осевого скелета, скелета головы (черепной коробки, предохраняющей головной мозг и связанной с жаберным и челюстным аппаратом), скелета грудного (плечевого) и тазового поясов и плавников – парных и непарных (рис. 8).
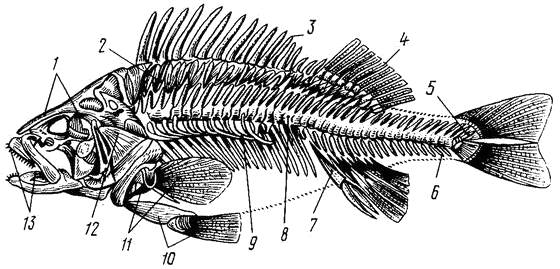
Рис. 8. Скелет костистой рыбы (окуня) (по Баклашовой, 1980):
1 – кости черепа, 2 – основные элементы спинного плавника, 3, 4 – лучи спинного плавника,
5 – последние позвонки, удерживающие хвостовой плавник, б – хвостовые позвонки,
7 – основные элементы анального плавника, 8 – туловищные позвонки, 9 – ребра с придатками,
10 – кости и лучи брюшного плавника, 11 – кости и лучи грудного плавника, 12 – жаберная крышка,
13 – верхняя и нижняя челюсти
Осевой скелет может быть представлен хордой или позвоночником. У круглоротых, осетровых и двоякодышащих хорда сохраняется в течение всей жизни. У всех остальных рыб хорда имеется на ранних этапах развития, а у взрослых заменяется позвоночником, состоящим из позвонков.
Черепная коробка соединена с позвоночником неподвижно. Шеи у рыб нет. Это вызывается спецификой образа жизни и среды обитания – необходимостью разрезать головой воду.
В процессе эволюции происходило усложнение и окостенение скелета. У круглоротых хорда тянется от задней части черепа до хвоста в виде цельного несегментированного тяжа, состоящего из хрящевыхи соединительнотканных элементов (спинная струна), к которому сверху плотно прилегают хрящевые позвоночные дуги. Хорда осетровых также ещё не дифференцирована. У пластиножаберных (акуловых) рыб охрящевевшая оболочка хорды образует амфицельные (двояковогнутые) позвонки.
Костистые рыбы имеют уже окостеневший позвоночник. В нём выделяют туловищный и хвостовой отделы. Туловищный отдел расчленен на типичные позвонки – амфицельные, в которых различают тело, верхнюю дугу с верхними (невральными) остистыми отростками (защищающими спинной мозг) и большие нижние дуги с нижними отростками. В туловищном отделе к позвоночнику (к поперечным отросткам или к телу позвонка) прикрепляются ребра. В хвостовом отделе поперечные отростки, смыкаясь, образуют нижнюю (гемальную) дугу, которая оканчивается нижним остистым отростком. В гемальном канале проходят хвостовые артерия и вена. Последний хвостовой позвонок уплощён и служит для прикрепления лучей хвостового плавника; он часто меняет обычную форму: удлиняется и загибается концом вверх, образуя уростиль.
Количество позвонков определяется рядом внутренних и внешних факторов и служит систематическим признаком рыбы. Например, у северной сельди их 57, речного угря – 114, сома – 72, луны–рыбы – 17, судака – 44. В пределах вида известна зависимость количества позвонков (и лучей в грудном и анальном плавниках) от температуры: повышение температуры в период эмбриогенеза вызывает уменьшение их числа.
Кроме ребер, опорную функцию у костистых рыб выполняют тонкие ‛мускульные‛ – межмышечные, или ‛туловищные‛, косточки, пронизывающие мышцы. Эти косточки образованы окостеневшими сухожилиями. Больше всего их у карповых рыб.
Череп рыб, так же как и осевой скелет, постепенно усложняется в процессе эволюции. У круглоротых нет черепной коробки; у них имеются отдельные, связанные соединительной тканью, хрящевые мозговые капсулы (обонятельная, слуховая, глазная).
Хрящевые рыбы (акулы, скаты) имеют уже хрящевой череп, образованный сросшимися мозговыми капсулами, челюсти, вооруженные зубами. Челюстной аппарат соединяется с черепом. У осетровых, кроме того, возникает ряд кожных накладных костей, прикрывающих череп сверху, и настоящих костей, ограждающих череп снизу.
Высшие костистые рыбы в эмбриональном периоде проходят все эти этапы: сначала у них закладывается и развивается хрящевой череп, позднее происходит окостенение его, причем наряду с замещающими костями возникают и покровные.
В черепе костистых рыб выделяют два отдела: мозговой (осевой) и висцеральный (рис. 9). Многочисленные кости осевого отдела соединены неподвижно. Этим достигается надежная защита мозга. Висцеральный отдел черепа образуется челюстным и жаберным аппаратом. Он состоит из челюстной, подъязычной и пяти жаберных дуг, прикрытых жаберной крышкой.
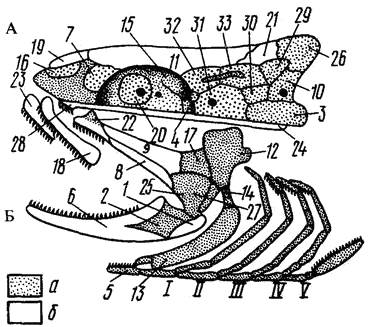
Рис. 9. Расположение костей в черепе костистой рыбы:
А – висцеральный скелет; Б – мозговой; жаберная крышка не нарисована
(по Наумову, Карташеву, 1979):
а – основные кости и хрящ, б – покровные кости;
1 – угловая, 2 – сочленовная, 3 – основная затылочная, 4 – основная клиновидная, 5 – копула,
6 - зубная, 7 – боковая обонятельная, 8 – наружная крыловидная, 9 – внутренняя крыловидная,
10 – боковая затылочная, 11 – лобная; 12 – подвесок, 13 – гиоид, 14 – окостеневшая связка,
15 – боковая клиновидная, 16 – средняя обонятельная, 17 – задняя крыловидная,
18 – верхнечелюстная, 19 – носовая, 20 – глазоклиновидная, 21 – теменная, 22 – небная,
23 – предчелюстная, 24 – парасфеноид, 25 – квадратная, 26 – верхняя затылочная,
27 – дополнительная, 28 – сошник, 29–30 – ушные кости, I – IV – жаберные дуги
Жаберная крышка состоит из четырех костей (рис. 10): предкрышечной – ргаеoperculum, крышечной – operculum, межкрышечной – interoperculum и подкрышечной – suboperculum. Они прикрывают пять жаберных дуг. На внутренней стороне четырех дуг расположены жаберные тычинки, а на внешней – жаберные лепестки (органы дыхания). Кости челюстного и жаберного аппарата сочленованы с черепом подвижно и, будучи взаимосвязанными, двигаются согласованно при помощи мышц.
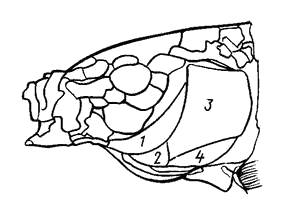
Рис. 10. Кости жаберной крышки:
1 – предкрышка, 2 – межкрышка, 3 – крышка, 4 – подкрышка
На 5-й жаберной дуге нет лепестков, у некоторых рыб эта дуга превращается в нижнеглоточную кость, и на ней образуются глоточные зубы, которые достигают наибольшего развития у карповых. Глоточные зубы располагаются в 1–3 ряда. Форма и расположение глоточных зубов являются систематическими признаками.
У карповых имеется также жерновок – роговое подушкообразное образование в верхней части глотки, служащее вместе с глоточными зубами для перетирания пищи.
Грудной (плечевой) пояс состоит из трех основных костей: ключицы – cleitrum, лопатки – scapula и коракоида – coracoid. К нему прикрепляются грудные плавники. Он сочленен при помощи задневисочной кости с черепом.
Тазовый пояс представлен у костистых рыб двумя срастающимися косточками, к которым прикрепляются лучи брюшных плавников. Он лежит в мышцах автономно, поэтому у некоторых видов может перемещаться далеко вперед, даже на горло, а иногда и вовсе исчезать. Скелет плавников являет собой опору, позволяющую рыбе пользоваться плавниками как рычагом или килем. У костистых рыб он представлен костными лучами, растягивающими кожистую плавательную перепонку.
Различают лучи жесткие и мягкие (нечленистые и членистые), которые, в свою очередь, разделяются на разветвленные и неразветвленные. Лучи соединяются с поясами конечностей или при помощи специальных косточек (грудной плавник), или непосредственно (брюшной плавник). Количество лучей в плавниках D и А подсчитывают при определении рыб, так как у костистых оно соответствует числу позвонков на определённом участке тела,
В соответствии с формой и строением различают несколько типов хвостового плавника (риc. 11).
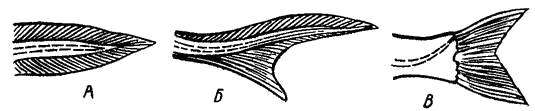
Рис. 11. Различные типы хвоста.
А – симметричный; Б – несимметричный (гетероцеркальный);
В – ложносимметричный (по Никольскому, 1974)
Все рыбы в эмбриональном и личиночном состоянии имеют прозрачную симметричную плавниковую кайму, огибающую конец позвоночника. Однако такая кайма у взрослых особей сохраняется только у круглоротых. У двоякодышащих позвоночник делит хвост на две равные части. Форма первично-симметричного хвоста (когда длинная ось тела делит его пополам) встречается редко.
У древних групп рыб (акулы, осетровые) хвост несимметричный, неравнолопастный или гетероцеркальный: конец позвоночника резко загибается вверх, лопасти хвоста разные.
У большинства костистых рыб (в том числе и у карпа) имеет место внутренняя асимметрия хвоста, образованная загнутым вверх уростилем, но при этом как вторичное явление развивается наружно-симметричная его форма. Такая форма называется ложно-симметричной.
Осевой скелет и скелет поясов выполняют опорную функцию, кроме того, к ним прикрепляются двигательные мышцы.
Дата добавления: 2015-06-22; просмотров: 1967;