Инфицирование бобового растения клубеньковыми бактериями
Для обеспечения нормального процесса инфицирования корневой системы клубеньковыми бактериями необходимо наличие довольно большого количества жизнеспособных клеток бактерий в прикорневой зоне. Мнения исследователей в отношении количества клеток, необходимых для обеспечения процесса инокуляции, различны. Так, по данным американского ученого О. Аллена (1966), для инокуляции мелкосеменных растений требуется 500—1000 клеток, для инокуляции крупносеменных — не менее 70 000 клеток на 1 семя. По мнению австралийского исследователя Дж. Винцента (1966), в момент инокуляции на каждое семя должно приходиться по крайней мере несколько сотен жизнеспособных и активных клеток клубеньковых бактерий. Имеются данные, что в ткань корня могут внедряться и единичные клетки.
При развитии корневой системы бобового растения размножение клубеньковых бактерий на поверхности корня стимулируется выделениями корня. Продукты разрушения корневых чехликов и волосков играют также немаловажную роль в обеспечении клубеньковых бактерий подходящим субстратом.
В ризосфере бобового растения резко стимулируется развитие клубеньковых бактерий, для злаковых растений такого явления не наблюдается.
На поверхности корня имеется слой слизистого вещества (матрица), образующийся независимо от наличия в ризосфере бактерий. Этот слой хорошо виден при исследовании в светооптическом микроскопе (рис. 147). Клубеньковые бактерии после инокуляции обычно устремляются к этому слою и скапливаются в нем (рис. 148) вследствие стимуля-ционного эффекта корня, проявляющегося даже на расстоянии до 30 мм.
В этот период, предшествующий внедрению клубеньковых бактерий в ткань корня, бактерии в ризосфере чрезвычайно подвижны. В ранних работах, в которых для исследований использовался световой микроскоп, клубеньковым бактериям, находящимся в зоне ризосферы, было дано название швермеров (гони-дий или зооспор) — «роящихся». С помощью метода Фэреуса (1957)можно наблюдать образование чрезвычайно быстро движущихся колоний швермеров в области кончика корня и корневых волосков. Колонии швермеров существуют очень короткое время—менее суток.
О механизме проникновения клубеньковых бактерий в корень растения существует ряд гипотез. Наиболее интересные из них следующие. Авторы одной из гипотез утверждают, что клубеньковые бактерии проникают в корень через повреждения эпидер-мальной и коровой ткани (особенно в местах ответвления боковых корней). Эта гипотеза была выдвинута на основании исследований Бриля (1888), вызвавшего образование клубеньков у бобовых растений путем прокалывания корней иглой, погруженной предварительно в суспензию клубеньковых бактерий. Как частный случай такой путь внедрения вполне реален. Например, у арахиса клубеньки преимущественно располагаются в пазухах ответвлений корней, что наводит на мысль о проникновении клубеньковых бактерий в корень через разрывы при прорастании боковых корней.
Интересна и не лишена оснований гипотеза о проникновении клубеньковых бактерий в ткань корня через корневые волоски. Путь прохождения клубеньковых бактерий через корневые волоски признает большинство исследователей.
Очень убедительно предположение П. Дарта и Ф. Мерсера (1965) о том, что клубеньковые бактерии внедряются в корень в виде мелких (0,1—0,4 мкм) кокковидных клеток через промежутки (0,3—0,4 мкм) целлюлозной фибриллярной сети первичной оболочки корневых волосков. Электронно-микроскопические фотографии (рис. 149) поверхности корня, полученные методом реплик, и факт мельчания клеток клубеньковых бактерий в ризосфере бобовых растений подтверждают это положение.
Не исключено, что клубеньковые бактерии могут проникать в корень через эпидермальные клетки молодых верхушек корня. По мнению Пражмовского (1889), бактерии могут проникать в корень только через молодую клеточную оболочку (корневых волосков или эпи-дермальных клеток) и совершенно не способны преодолевать химически измененный или оп-робковевший слой коры. Этим можно объяснить, что клубеньки обычно развиваются на молодых участках главного корня и появляющихся боковых корнях.
В последнее время большую популярность получила ауксинная гипотеза. Авторы этой гипотезы считают, что клубеньковые бактерии проникают в корень благодаря стимуляции синтеза р-индолилуксусной кислоты (гете-роауксина) из триптофана, имеющегося всегда в корневых выделениях растений. С наличием гетероауксина связывается искривление корневых волосков, которое обычно наблюдается при инфицировании корневой системы клубеньковыми бактериями (рис. 150).
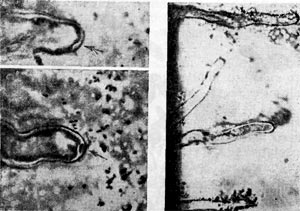
Рис. 148. Скопления клубеньковых бактерий вокруг корневых волосков. Стрелкой показано движение клеток клубеньковых бактерий к вершине корневого волоска клевера.
Источником бета-индолилуксусной кислоты в момент инфицирования растения, очевидно, служат не только растения, выделяющие через корневую систему триптофан, который многие виды бактерий, в том числе и клубеньковые, могут переводить в бета-индолилуксусную кислоту. Сами клубеньковые бактерии, а возможно, и другие виды почвенных микроорганизмов, живущие в зоне корня, также могут участвовать в синтезе гетероауксина.
Однако принимать безоговорочно ауксинную гипотезу нельзя. Действие гетероауксина неспецифично и вызывает искривление корневых волосков у разных видов растений, а не только бобовых. В то же время клубеньковые бактерии вызывают искривление корневых волосков лишь у бобовых растений, проявляя при этом довольно значительную избирательность. Если бы рассматриваемый эффект определялся только (З-индолилуксусной кислотой, то такой специфики не было бы. Кроме того, характер изменений корневых волосков под влиянием клубеньковых бактерий несколько иной, чем под влиянием гетероауксина.
Следует также отметить, что в отдельных случаях инфицированию подвергаются неискривленные корневые волоски. Наблюдения показывают, что у люцерны и гороха искривляются и закручиваются 60—70% корневых волосков, а у клевера — около 50%. У некоторых видов клевера эта реакция отмечается не более чем у 1/4 части заражаемых волосков. В реакции искривления, очевидно, имеет большое значение состояние корневого волоска. Растущие корневые волоски наиболее чувствительны к действию веществ, вырабатываемых бактериями.
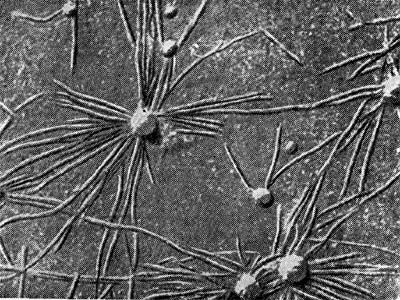
Рис. 149. Клетки клубеньковых бактерий на поверхности инфицированного корневого волоска бобового растения (по П. Дарту, Ф. Мерсеру).
Известно, что клубеньковые бактерии вызывают размягчение стенок корневых волосков. Однако ни целлюлазы, ни пектинолитических ферментов они не образуют. В связи с этим было высказано предположение, что клубеньковые бактерии проникают в корень благодаря выделению ими слизи полисахаридной природы, вызывающей синтез растениями фермента поли-галактуроназы. Этот фермент, разрушая пектиновые вещества, влияет на оболочку корневых волосков, делая ее более пластичной и проницаемой. В небольших количествах полигалактуроназа всегда присутствует в корневых волосках и, очевидно, вызывая частичное растворение соответствующих компонентов оболочки, позволяет клетке растягиваться.
Некоторые исследователи полагают, что клубеньковые бактерии проникают в корень благодаря бактериям-спутникам, продуцирующим пектинолитические ферменты. Эта гипотеза была выдвинута на основании следующих фактов. При микроскопировании корневых волосков многие исследователи отмечали наличие светлого пятна, около которого скапливаются клубеньковые бактерии. Это пятно, возможно, является признаком начала мацерации (разрушения) ткани протопектиназой по аналогии с таким же признаком, наблюдающимся у растений при многих бактериальных заболеваниях. Кроме того, установлено, что авирулентные культуры клубеньковых бактерий в присутствии бактерий, продуцирующих пектинолитические ферменты, становятся способными проникать в корень.
Следует отметить еще одну гипотезу, по которой клубеньковые бактерии попадают в корень при образовании пальцевидного впячивания поверхности корневого волоска. На электрон-нограмме среза корневого волоска, подтверждающей эту гипотезу (рис. 150, 5), виден изогнутый в виде ручки зонтика корневой волосок, в изгибе которого находится скопление клубеньковых бактерий. Клубеньковые бактерии как бы втягиваются (проглатываются) корневым волоском (подобно пиноцитозу).
Гипотеза инвагинации, по существу, не может быть отделена от ауксинной или ферментативной гипотезы, поскольку инвагинация происходит в результате воздействия либо ауксинного, либо ферментного фактора.
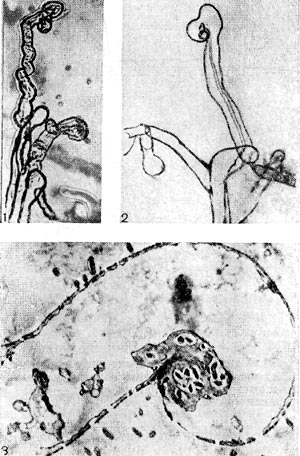
Рис. 150. Искривление корневых волосков бобовых растений в присутствии клубеньковых бактерий: 1,2 — увел, х 120; 3 — ультратонкий срез через корневой волосок. Увел. X 10 000 (по К. Сальману и Г. Фэреусу).
Процесс внедрения клубеньковых бактерий в ткань корня одинаков у всех видов бобовых растений и состоит из двух фаз. В первую фазу происходит инфицирование корневых волосков. Во вторую фазу интенсивно идет процесс образования клубеньков. Продолжительность фаз различна у разных видов растений: у Trifolium fragiferum первая фаза продолжается 6 дней, у Trifolium nigrescens — 3 дня. В некоторых случаях очень трудно обнаружить границы между фазами. Наиболее интенсивное внедрение клубеньковых бактерий в корневые волоски происходит на ранних этапах развития растения. Вторая фаза заканчивается в период массового образования клубеньков. Нередко внедрение клубеньковых бактерий в корневые волоски продолжается уже и после того, как клубеньки сформировались на корнях. Эта так называемая избыточная или дополнительная инфекция происходит потому, что инфицирование волосков не прекращается длительное время. В более поздние сроки заражения клубеньки обычно размещаются ниже по корню.
Тип развития, структура и плотность корневых волосков не влияют на скорость внедрения клубеньковых бактерий. Места образования клубеньков не всегда связаны с местами расположения инфицированных волосков.
Проникнув в корень (через корневой волосок, эпидермальную клетку, места повреждений корня), клубеньковые бактерии далее перемещаются в ткани корня растения. Наиболее легко бактерии проходят через межклеточные пространства.
Внедриться в ткань корня может или одиночная клетка, или группа клеток бактерий. Если внедрилась отдельная клетка, она и в дальнейшем может перемещаться по ткани как одиночка. Путь инфицирования корня одиночными клетками свойствен растениям люпина.
Однако в большинстве случаев внедрившаяся клетка, активно размножаясь, образует так называемые инфекционные нити (или инфекционные тджи) и уже в виде таких нитей перемещается в ткани растения.
Термин «инфекционная нить» возник на основе изучения процесса инфицирования в световом микроскопе. Начиная с работ Бейеринка, инфекционная нить стала рассматриваться как слизистая гифообразная масса с заключенными в нее размножающимися бактериями.
По существу, инфекционная нить — это колония размножившихся бактерий. Началом ее служит то место, куда проникла отдельная клетка или группа клеток. Не исключено, что колония бактерий (а следовательно, и будущая инфекционная нить) начинает формироваться еще на поверхности корня до момента внедрения бактерий в корень.
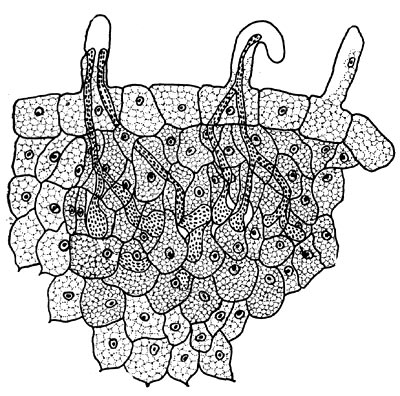
Рис. 151. Схема возникновения инфекционных нитей. В неискривленном корневом волоске видны две инфекционные нити, в искривленном — одна разветвляющаяся (по Ф. Бибердорфу).
Количество инфицированных корневых волосков значительно различается у отдельных растений. Обычно инфекционные нити появляются в деформированных, искривленных корневых волосках. Однако есть указания, что и в прямых волосках иногда обнаруживаются подобные нити. Чаще в корневых волосках наблюдается одна разветвляющаяся нить, реже две. В некоторых случаях в одном корневом волоске имеется несколько нитей или же в нескольких имеются общие нити заражения, дающие начало одному клубеньку (рис. 151).
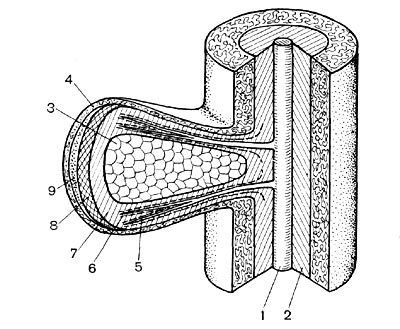
Рис. 152. Схема, показывающая связь корня с клубеньком: 1 — сосудистая ткань корня; 2 — корневая эндодерма; 3 — бактёроидная зона клубенька; 4 — кора клубенька; 5 — сосудистые пучки клубенька; б — эндодерма сосудистых пучков; 7 — клубеньковая эндодерма; 8 — меристема; 9 — верхушечная меристема.
Процент инфицированных корневых волосков в общем количестве деформированных необъяснимо низок. Он обычно колеблется от 0,6 до 3,2, изредка достигая 8,0. Доля удачных инфекций еще ниже, поскольку среди инфекционных нитей имеется много (до 80%) так называемых абортивных нитей, прекративших свое развитие. Скорость продвижения нормально развивающихся инфекционных нитей в растении — 5—8 мкм в час. При такой скорости путь через корневой волосок длиной 100—200 мкм инфекционная нить может пройти в течение одних суток.
Дата добавления: 2015-06-12; просмотров: 1620;