Систематика спорообразующих анаэробных бактерий
В настоящее время имеются три наиболее известные системы классификации анаэробов: Н. А. Красильникова (1949), Берги (1957) и Прево (1957, 1967). По Красильникову, все спорообразующие бактерии объединяются в семейство ВасШасеае. Это семейство имеет два рода — Bacillus и Clostridium. В род Bacillus входят как аэробные, так и анаэробные бактерии (последние составляют Х-группу), которые не образуют гранулезы и не раздуваются (либо слабо раздуваются) при спорообразовании. Род Clostridium объединяет бактерии, клетки которых в процессе спорообразования раздуваются с приобретением клостридиальной (лимоновидной) формы и накапливают гранулезу.
За прошедшие 20 лет после издания определителя Красильникова накопилось много данных об анаэробных микроорганизмах, их видовом разнообразии, биохимии, физиологии и цитологии. Были обнаружены новые физиологические группы, относящиеся к разным родам, и выделено много новых видов спорообразующих анаэробов.
Определитель Прево, Турпина и Кайзера наиболее полно описывает анаэробные микроорганизмы, известные в настоящее время микробиологам. Около 200 видов спорообразующих анаэробных бактерий распределяются в нем по трем порядкам: Clostridiales, Plectridiales и Sporovibrionales (схема 3 на табл. 36).
Морфологические различия между вегетативными клетками различных видов очень незначительны, а спорулирующие клетки и сами споры необычайно разнообразны. Это явление имеет очень большое значение для разделения анаэробов на систематические группы. Для разделения на порядки служит форма вегетативных клеток (прямые или изогнутые палочки) и расположение спор в спорангиях (центральное, субтерминальное и терминальное). При разделении на семейства и роды принимаются во внимание грамокраска, подвижность клеток, способность образовывать капсулу. Различные сочетания этих признаков позволили разделить все спорообразующие анаэробные бактерии на 5 семейств и 10 родов.
Порядок Clostridiales. К нему относятся подвижные и неподвижные грамположительные и грамотрицательные палочки, раздувающиеся при спорообразовании. Споры расположены центрально и субтерминально. Имеются пигментированные виды. В основном анаэробные, но возможны факультативные формы. В порядок входят два семейства.
Семейство Endosporaceae включает грамотрицательные клостридии; споры расположены центрально или субтерминально. В этом семействе два рода: Endosporus — подвижные формы и Paraplectrum — неподвижные, образуют капсулу. Семейство Clostridiaceae включает грамположительные клостридии. Споры центральные или субтерминальные. Подразделяется на три рода: Inflabilis — неподвижные формы, Welchia — неподвижные капсулированные палочки, Clostridium — подвижные формы.
Порядок Plectridiales. Подвижные и неподвижные, грамположительные и грамотрицательные палочки. Споры образуются терминально. Порядок подразделяется на два семейства.
Семейство Terminosporaceae — грамотрицательные плектридии. Семейство состоит из двух родов: Terminosporus — подвижные и Cadu-ceus — неподвижные формы.
Семейство Plectridiaceae — грамположительные плектридии. В этом семействе два рода: Plectridium — подвижные и Acuformis — неподвижные организмы.
Порядок Sporovibrionales включает спорообразующие подвижные анаэробные бактерии с изогнутыми (вибрионоподобными) грамотрицательными клетками. Большинство из них способно восстанавливать сульфаты и нитраты.
В этом порядке одно семейство — Sporovibrionaceae. В него входит один род — Sporovibrio, объединяющий четыре вида спорообразующих анаэробов. Один из этих видов — Sporovibrio ferrooxydans — не восстанавливает сульфатов, а железо окисляет в гидрат железа. По Кэмпбеллу и Постгейту, все спорообразующие анаэробы, обладающие постоянной способностью восстанавливать сульфаты, были выделены в новый род — Desulfo-tomaculum. В него входят облигатные анаэробы с грамотрицательными, прямыми или изогнутыми палочками, раздувающимися у термофильных форм. Споры формируются терминально или субтерминально. Бактерии подвижные, жгутикование перитрихиальное, восстанавливают сульфаты до сульфидов. Обладают цито-хромами (класс протогеминов). Состав ДНК колеблется в пределах 41,7—49,2 мол.% Г+Ц.
Рис. 64. Сульфатредуцирующая спороносная бактерия. Клетки при спорообразовании раздуваются. Увел. Х2700.
Неспоровые сульфатредуцирующие бактерии из рода Desulfovibrio четко отличаются от спорообразующих по проценту Г + Ц в ДНК, наличию пигмента десульфовиридина и цито-хрома, монотрихиальным жгутикованием и отсутствием спорообразования.
Род Desulfotomaculum включает четыре вида. D. nigrifleans — типовой вид. Споры овоидные, субтерминальные. Бактерии могут расти без сульфатов на среде с пируватом. Глюкозу и другие углеводы не используют, термофильные. Температурный оптимум 55° С. Процент Г + Ц в ДНК 49,2. D. ruminis. Споры овоидные, терминальные или субтерминальные. Бактерии способны расти без сульфатов на среде с пируватом. Глюкозу и другие углеводы не сбраживают, мезофилы, Г + Ц в ДНК-49,2%.
D. orientis. Споры круглые, могут располагаться в клетках центрально, эксцентрально и терминально. Глюкозу и другие углеводы бактерии не сбраживают, не растут на средах с пируватом без сульфатов, мезофилы. Процент Г+Ц в ДНК 45,1. Японские авторы описали еще один вид спорообразующих сульфатредуцирующих бактерий — D. antarcticum. Споры этих бактерий овоидные, располагаются в клетках центрально или терминально. Нуклеотидный состав ДНК неизвестен. Этот вид отличается от других видов рода Desulfotomaculum способностью использовать глюкозу и разжижать желатин.
Аэробные спрообразующие бактерии. Род бациллюс (BACILLUS)
Аэробные спорообразующие бактерии составляют довольно обширную группу микроорганизмов. Они широко распространены в природе и играют большую роль в разнообразных биологических процессах. С использованием этих бактерий в промышленности освоено производство ценных ферментов, антибиотиков, органических кислот и других соединений.
Среди этой группы бактерий имеются и патогенные для человека и животных формы. В этом отношении особенно тщательно изучен сибиреязвенный бацилл. Имеются данные о фитопатогенных свойствах отдельных видов спорообразующих бактерий, обусловливающих развитие гнили и других поражений у растений.
Некоторые виды спорообразующих бактерий являются возбудителями болезней полезных насекомых. Образуя единственные в своем роде покоящиеся формы организмов — споры, эти бактерии служат объектом многочисленных исследований для выяснения природы ана- и парабио-тического состояния живой материи. С другой стороны, благодаря образованию спор, обладающих среди известных форм живых организмов наиболее высокой устойчивостью к неблагоприятным воздействиям внешней среды, спорообразующие бактерии представляют собой основной фактор биологического разрушения пищевых продуктов и различных других материалов.
Борьба со спорообразующими бактериями является одной из главных практических задач в консервной промышленности, при консервации крови и многих других процессах хранения и переработки сельскохозяйственных и пищевых продуктов.
Спорообразующие бактерии распространены повсеместно — в воздухе, водоемах, на растительных и животных остатках и других естественных субстратах.
Благодаря способности образовывать споры, обладающие большой устойчивостью к внешним воздействиям, спорообразующие бактерии остаются жизнеспособными при самых неблагоприятных условиях. Эти формы микроорганизмов выдерживают резкие колебания температуры, отсутствие влаги и воздуха, действие различных химических соединений, обычно губительно влияющих на живые организмы. Поэтому среди бесчисленного множества самых различных форм микроорганизмов спорообразующие бактерии являются наиболее устойчивыми и приспособленными к изменениям среды. Часто при анализе разных субстратов, подвергшихся неблагоприятным воздействиям, единственными представителями микробов бывают зародыши спороносных бактерий — споры.
Таким образом, спороносные бактерии имеют по сравнению с другими формами микробов большие возможности приспосабливаться к тем или другим условиям среды.
Спорообразующие бактерии распространены в морях и океанах, включая их глубокие слои и арктические воды. Работами А. Е. К рисса, С. И. Кузнецова и других советских ученых установлено распространение разнообразных видов спорообразующих бактерий в водоемах и их участие во многих биологических процессах среды своего обитания.
Большинство видов спорообразующих бактерий, подобно другим микроорганизмам, в основном обитает в почве. Однако не все почвы богаты спороносными бактериями. Содержание их в одной и той же почве различно в зависимости от окультуренности почвы, метеорологических, сезонных изменений и многих других факторов.
Почвы разных типов или возраста содержат неодинаковое число спорообразующих бактерий. Значительная часть таких бактерий обитает в бурых, сероземных, пустынных почвах, характеризующихся малым содержанием органического вещества. В богатых перегноем почвах спорообразующих бактерий сравнительно мало, например в арктических, дерново-подзолистых, лесных и горнолуговых.
Наряду с изменениями в общем (валовом) содержании спорообразующих бактерий особенно большие различия отмечаются в их видовом составе. В одних типах почв преобладают одни виды, в других — другие.
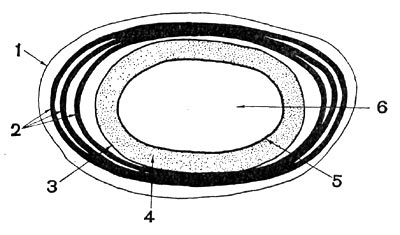
Рис. 65. Схематическое изображение строения споры: 1 — экзоспориум; 2 — слои споровой оболочки; 3 — внешняя мембрана споры; 4 — кора; 5 — внутренняя мембрана споры; 6 — сердцевина.
Фундаментальными работами в области экологии аэробных спорообразующих бактерий явились исследования Е. Н. Мишустина и его сотрудников. Были выявлены определенные закономерности распространения в почве отдельных видов спороносных бактерий. Так, установлено, что в биологических процессах, протекающих в почве, неспороносные бактерии связаны с превращениями гумуса на первых стадиях его минерализации, и поэтому по соотношению числа спороносных и неспороносных бактерий можно судить об интенсивности минерализации органического вещества в почве. Были выявлены характерные группировки спорообразующих бактерий, свойственные определенным почвенным типам и эколого-геогра-фическим зонам. На основе этих работ развились представления о показательных свойствах спорообразующих бактерий, отражающих почвообразовательный процесс, и описаны доминантные и типичные группы этих организмов для отдельных почвенно-климатических зон.
Закономерности эколого-географического распространения в почве различных видов споро-образующих бактерий достаточно рельефно обнаруживаются и по вертикальной зональности — при обследовании горных районов с большим разнообразием типов почвы, растительных формаций и климатических условий. С увеличением высоты в горах содержание и состав бактерий претерпевают закономерные изменения, в общем подобные тем, которые отмечаются при обследовании разных типов почв по широтной зональности.
Таким образом, изучение эколого-географического распространения спорообразующих бактерий выявляет специфику микробного населения различных типов почв и подтверждает учение В. В. Докучаева о зонах природы, горизонтальной и вертикальной зональности почвенного покрова. Общее количество и видовой состав споро-образующих бактерий в почве подвержены большим изменениям в зависимости от ее влажности и температуры.
При повышении влажности почвы после обильного дождя отмечается интенсивное размножение неспороносных бактерий. Их массовое развитие приводит к угнетению роста и развития спороносных бактерий. Их количество в почве может уменьшиться. Такое явление наблюдается при влажности почвы в пределах 50—60% и более от полной влаго-емкости. При умеренной влажности, в особенности в несколько подсохшей почве, количество спороносных бактерий увеличивается, а число неспороносных бактерий значительно уменьшается.
Много разнообразных спорообразующих бактерий обнаруживается в почвах со сравнительно высоким температурным режимом. Обильны бактериями почвы южных районов, а на севере и в почвах полярного круга их сравнительно мало. Значительный перегрев почвы на юге губительно действует на неспороносные бактерии, в то время как споровые формы переносят его хорошо.
У хорошо известного вида спорообразующих бактерий — Вас. mycoides установлено наличие эколого-географических разновидностей, характерных для отдельных почвенно-климатиче-ских зон. Выявлено, что у культур этих бактерий с удалением от севера к югу положение оптимальной и максимальной температур развития соответственно повышается. Южные культуры бактерий интенсивнее размножаются и обладают гораздо более высоким осмотическим давлением в клетках, нежели северные. У других видов спороносных бактерий такой приспособительной реакции к температурным условиям среды не отмечено.
Своеобразие эколого-географических условий приводит к расселению отдельных видов и разновидностей спорообразующих бактерий в определенных типах почв. Так, установлено очаговое распространение в красноземах неестественных, правозавитых, или так называемых инверсивных, форм Вас. mycoides. В солончаках обнаружены особые, галофильные разновидности спорообразующих бактерий, приспособившиеся к жизни в среде с высокой концентрацией солей. На питательных средах с небольшими концентрациями минеральных солей эти культуры развиваются очень слабо. Растительный покров оказывает значительное влияние на количественный и качественный состав спорообразующих бактерий в почве.
В почве, непосредственно прилегающей к корням растений (ризосфера), основная масса микробов представлена неспороносными бактериями. Споровых форм здесь очень мало. Однако на разлагающихся растительных остатках, которых много в почве осенью, спорооб-разующие бактерии обнаруживаются в значительном количестве.
Многие виды спорообразующих бактерий активно продуцируют ферменты, разлагающие обычно трудно разрушающиеся растительные остатки. Благодаря деятельности микроорганизмов почва обогащается ценными питательными веществами и освобождается от многих вредных организмов и продуктов. Возделывание в течение ряда лет одной и той же культуры может значительно изменить качественный состав спорообразующих бактерий в почве. Сдвиги в видовом составе споровых бактерий отмечаются при многолетнем культивировании люцерны и хлопчатника. В севооборотах эти различия несколько стираются, однако для микробиологических исследований как споровых, так и других форм бактерий в разных почвах знание истории поля обязательно. Накопление в почве спорообразующих бактерий-антагонистов оказывает благотворное действие на ее оздоровление, приводя к снижению в почве количества болезнетворных микробов.
Установлена ведущая роль спороносных бактерий в более глубоких стадиях распада веществ в компостах и других органических, соединений почвы. Обработка почв и агрохимические мероприятия, приводящие к уменьшению содержания подвижного органического вещества в почве, сопровождаются повышением содержания бациллярной флоры. Подобное явление отмечается и при длительном использовании минеральных удобрений. Количество спорообразующих бактерий повышается в нижних слоях почвы.
Сравнительно высокое содержание в почве спор бактерий является отражением определенной стадии в развитии этих организмов и не может свидетельствовать о неактивности спороносных бактерий.
О спорах бактерий, их химическом составе и структуре, процессах образования и прорастания подробно было рассказано в предыдущей главе. Здесь же мы остановимся на отличительных чертах, характерных для аэробных спорообразующих бактерий.
Поверхность наружной оболочки спор разнообразна у аэробных бактерий: либо гладкая, либо с выростами, выступами и почками. Оболочка составляет значительную часть споры. Основными ее компонентами являются белки (60—90%) и липиды. Состав аминокислот в белках оболочки неодинаков у бактериальных спор разных видов, однако большая часть цистина, содержащегося в значительном количестве в спорах, обнаруживается именно в оболочке.
Под оболочкой располагается кора споры (рис. 65), состоящая в основном из муреина. который является нерастворимым структурным материалом коры.
В течение многих лет покоящаяся спора рассматривалась как почти инертное тело, обладающее несколькими ферментами или вовсе лишенное их. Некоторых ферментных систем, имеющихся в вегетативной клетке, например цитохромов и ферментов цикла трикарбоновых кислот, ничтожно мало в споре. Это указывает на то, что в спорах аэробный энергетический метаболизм осуществляется иным путем, чем в клетках. Аналогичные ферменты спор и вегетативных клеток в спорах более термоустойчивы.
Споры аэробных бактерий обладают различной термоустойчивостью, что обусловлено как видовыми особенностями, так и условиями их образования. Установлено, что споры, образующиеся при высокой температуре, а также полученные на средах с высоким содержанием кальция, парааминобензойной кислоты и некоторых жирных кислот, более жароустойчивы.
Споры более устойчивы, чем вегетативные клетки, от которых они образовались, к воздействию проникающей радиации, ультразвука, гидростатического давления, замораживания, разрежения и др. Устойчивость спор к кислотам и щелочам во многом зависит от их принадлежности к различным видам бактерий. Споры некоторых культур бактерий остаются жизнеспособными даже при кипячении в концентрированной соляной кислоте в течение 20 мин. При этом резистентность спор во много раз повышается при их обезвоживании.
К спиртам и другим органическим растворителям устойчивость спор намного выше, чем у вегетативных клеток. Споры менее устойчивы к действию алкилирующих агентов. При воздействии комплекса определенных ферментов удается достигнуть растворения спор.
Механизм устойчивости спор к действию высокой температуры, проникающей радиации и других физико-химических факторов полностью еще не выяснен. Термоустойчивость спор объясняется наличием кальция, дипиколиновой кислоты и в некоторой степени диаминопимелиновой кислоты. Эти вещества определяют в значительной мере и резистентность спор к действию радиации. Полагают, что споры содержат комплекс особых веществ, подобных хелатным соединениям, которые предохраняют жизненно важные структурные элементы от губительного воздействия физико-химических факторов. Важным фактором, приводящим к образованию спор, является отсутствие для данного вида аэробных бактерий благоприятных условий для роста и развития. Способствует спорообразованию обеднение питательной среды.
У аэробных бактерий анаэробные условия выращивания подавляют как спорообразование, так и рост. Спорообразование аэробных бактерий протекает лишь в условиях доступа кислорода. Интенсивность споруляции подавляется в разной степени при выращивании бактерий в неблагоприятных условиях — при температуре выше оптимальной или реакции среды менее подходящей для обильного развития. Неблагоприятные условия роста, ускоряющие споруляцию бактерий, как правило, не обусловливают обильного «урожая» спор.
Разные виды аэробных бактерий отличаются расположением и формой спор. Эти признаки имеют важное систематическое значение. У некоторых видов расположение спор строго определенное — центральное, околоконцевое и концевое. У других видов не наблюдается строгой локализации. Высокое содержание в питательной среде источников азотного и углеродного питания, обеспечивающих максимальный рост бактерий, может подавлять индукцию спорообразования. Так, потребность в глюкозе — основном энергетическом источнике вегетативного роста аэробных бактерий — является несущественной для спорообразования некоторых видов, а в отдельных случаях она угнетает процесс споруляции. Культуры определенных видов бактерий более интенсивно спорулируют при уменьшении в среде аланина, валина, лейцина и изо-лейцина. У других бактерий отсутствие в среде лейцина и серусодержащих аминокислот угнетает спорообразование.
Интенсивность процесса образования спор в значительной мере обусловлена наличием в среде минеральных солей, содержащих ионы кальция, магния, калия, марганца, железа, цинка, меди и некоторых редких металлов (кобальта, лития, кадмия, никеля). Оптимальная концентрация минеральных солей и металлов различна в зависимости от вида выращиваемой культуры бактерий, условий ее роста. Роль минеральных солей и отдельных ионов металлов настолько велика, что в их отсутствии спороношение может вовсе не наблюдаться. Изменения концентраций их в питательной среде могут оказать иногда резко различное действие.
Спорообразование отдельных культур бактерий настолько зависит от наличия минимальных концентраций некоторых металлов, что эта особенность используется в аналитических целях.
Роль минеральных солей различных металлов в образовании бактериальных спор установлена недавно, и многие вопросы механизма их действия пока не выяснены. Полагают, что их специфическое действие связано с активированием различных ферментативных систем. Значение таких элементов, как кальций, заключается также и в том, что они способствуют образованию разнообразных низкомолекулярных веществ в спорах, обеспечивающих их термостабильность.
Сравнительно слабо изучено влияние на образование спор различных витаминов и других ростовых веществ. Некоторое усиление этих процессов отмечено под влиянием параамино-бензойной и фолиевой кислот.
В результате всестороннего изучения биохимии спорообразования показано, что при спору-ляции не образуются новые ферментные системы. Аналогичные ферменты, обнаруживаемые в вегетативной клетке и споре, имеют одинаковые молекулярный вес, аминокислотный состав и другие физико-химические свойства. Теплоустойчивость ферментов спор вызвана присутствием в них ионов кальция, марганца и других металлов. Данные исследований свидетельствуют о том, что природа и функция ферментативного аппарата, осуществляющего синтез белков в споре и материнской вегетативной клетке, одинаковы. Основной системой транспорта электронов в спорах является растворимая флавопротеино-ксидаза. При споруляции выявлена высокая активность ряда ферментов, например аргиназы, некоторых нуклеаз. Основная масса белков споры синтезируется заново в период споруляции. Одновременно отмечается процесс распада белков, причем многими авторами обнаружена тесная взаимосвязь протеолитической активности с функцией спорообразования у бактерий. Из вновь синтезированных белков лишь незначительная часть используется для создания структурных элементов споры. В целом споруляцию можно охарактеризовать как процесс интенсивного распада белков и всех основных типов РНК материнской клетки с одновременным образованием новых белковых компонентов.
Различия в составе белков споры и вегетативной клетки выявляются серологическими методами. По антигенной структуре спорулирующая и аспорогенная формы культур одного и того же вида различны. Не установлено существование какого-либо антигена, общего для спор бактерий разных видов. Наоборот, по антигенной структуре имеется достаточно отчетливая дифференциация между спорами разных видов.
В способе прорастания споры и продолжительности процесса также отмечается определенное постоянство, характерное для различных видов бактерий. Старые споры прорастают в течение более длительного периода, чем молодые. Время прорастания молодых спор является довольно постоянным у различных бактерий и может служить видовым признаком. Если перенести только что сформировавшиеся споры в благоприятную питательную среду, то прорастание их начинается не сразу, а по истечении 2—3 ч.
Установлено, что на начальной стадии прорастания спор происходит выделение в среду так называемого экссудата прорастания, характеризующегося специфическим поглощением ультрафиолетовых лучей. Вещество это было выделено и идентифицировано как кальциевая соль дипиколиновой кислоты (пиридин — 2,6-дикарбоксиловая кислота). Дипиколинат кальция составляет приблизительно половину сухой массы экссудата прорастания, что соответствует 15% от общей сухой массы покоящихся спор. Остальная часть экссудата прорастания представлена свободными аминокислотами, высокомолекулярными и низкомолекулярными белковыми веществами. Обнаружено особенно большое количество недиализируемого гексозаминсодержащего пептида с молекулярным весом порядка 10 000, в состав которого входят диаминопимелиновая кислота, аланин, глюта-миновая кислота и гексозамины. Условно это соединение названо ДАП-гексозаминпептид. Это соединение входит в состав оболочки спор, в отличие от дипиколиновой кислоты, обнаруживаемой внутри спор.
Среди факторов, способствующих процессу прорастания спор, большое внимание уделяется тепловой активации, действие которой более выражено при наличии в среде некоторых химических соединений. Тепловая активация достигается предварительным кипячением суспензии спор при 97—100° С в течение 10—15 мин или при 65° С в течение 2 ч. Иногда этот эффект достигается при обычной пастеризации. Тепловая активация прорастания спор носит общий характер для разных видов спороносных бактерий, хотя она более специфична для аэробов, чем анаэробных видов. Обычно тепловое воздействие усиливает и ускоряет прорастание спор в течение короткого промежутка времени. Действие подобной обработки спор, как правило, преходяще, хотя при этом отмечается и некоторое изменение условий, необходимых для течения процесса прорастания спор. Так, для лучшего прорастания спор сенного бацилла без предварительной тепловой обработки необходимо присутствие в среде L-аланина, аденозина, глюкозы и тирозина, тогда как после тепловой обработки оптимальные условия прорастания спор обеспечивают наличие лишь глюкозы и аденозина. Установлено также, что сохранение в течение длительного времени спор при 37° С после их тепловой активации ведет к деактивации. Тепловая обработка этих спор после длительного хранения вновь ведет к реактивации прорастания.
Имеются данные о некотором активирующем влиянии на прорастание спор умеренного воздействия ультразвука. Исследования последних лет показали, что интенсивность прорастания спор специфически повышается в присутствии некоторых веществ, в первую очередь L-аланина, пуриновых нуклеозидов и глюкозы. Иногда необходимо наличие, помимо L-аланина, ряда других аминокислот. Из рибозидов наиболее эффективным оказался аденозин.
Алании является наиболее специфическим активатором процесса прорастания спор из всех известных соединений, обладающих подобным действием. Оптимальные условия для прорастания спор анаэробных бактерий несколько более сложны, хотя факторы, активирующие процессы прорастания спор аэробных видов, и в этом случае оказывают активирующее действие.
Наряду с активаторами выявлен ряд веществ, угнетающих процесс прорастания спор. Прорастание спор у аэробных видов угнетается в присутствии оксина, некоторых мышьяковистых соединений, углекислого натрия, высоких концентраций ряда аминокислот — глицина, метионина, цистеина и валина. Характерно угнетающее действие D-аланина в отличие от L-аланина.
Механизм активирующего и угнетающего действия ряда факторов на прорастание спор остается неясным. Одни авторы приписывают активаторам определенную роль в процессах деполимеризации различных составных элементов в спорах, высвобождении и активации ряда ферментных систем, обусловливающих начало прорастания спор. Другие считают их энергетически активными соединениями, стимулирующими биохимические процессы в момент прорастания спор.
Обычно не все споры, вносимые в благоприятную среду, прорастают сразу. Некоторое их количество остается непроросшими в течение различного времени, не теряя, однако, своей жизнеспособности. Установлено, что при длительном хранении процент прорастающих спор постепенно снижается. Причина этого явления и природа подобных «дремлющих» спор остаются неясными.
В покоящейся споре имеются ферментные системы, выявляемые лишь после их активации под влиянием определенных факторов и химических соединений, которые, однако, исключают прорастание спор. К таким системам относятся прежде всего окислительные ферменты.
Активация их достигается обработкой спор незначительными количествами L-аланина, аденозина и некоторыми другими веществами, а также кипячением или пастеризацией суспензий спор. Перевод неактивной ферментной системы в активную без признаков прорастания спор считается первым этапом биохимических изменений в спорах, ведущих в дальнейшем к началу процесса их прорастания. Полагают, что под действием активаторов дипиколиновая кислота, связывающая окислительные и другие ферменты, подвергается деполимеризации с освобождением ферментных систем.
С помощью высокочувствительного микрореспирометра у бактериальных спор установлена дыхательная активность.
При изучении химического состава клеточной стенки спор различных видов аэробных бактерий обнаружены характерные различия в содержании ДАП-гексозаминпептида и других веществ — полисахаридов, липидов. ДАП-гек-созаминпептид высвобождается из стенки спор под действием литического агента, уподобляемого по своим свойствам лизоциму.
Дата добавления: 2015-06-12; просмотров: 1582;