Синтез жирных кислот
По сравнению с гликогеном жиры представляют более компактную форму хранения энергии, поскольку они менее окислены и гидратированы. При этом количество энергии, резервированное в виде нейтральных липидов в жировых клетках, ничем не ограничивается в отличие от гликогена. Центральным процессов в липогенеза является синтез жирных кислот, поскольку они входят в состав практически всех групп липидов. Кроме того, следует помнить, что основным источником энергии в жирах, способным трансформироваться в химическую энергию молекул АТФ, являются процессы окислительных превращений именно жирных кислот.
Общая характеристика биосинтеза жирных кислот:
1.Жирные кислоты могут синтезироваться из углеводов пищи через пируват или из аминокислот (при их избыточном поступлении) и накапливаются в виде триацилглицеролов
2. Основное место синтеза – печень. Кроме того, жирные кислоты синтезируются во многих тканях: почки, мозг, молочная железа, жировая ткань.
3.Ферменты синтеза локализованы в цитозоле клеток в отличие от ферментов окисления жирных кислот, которые находятся в митохондриях.
4.Синтез жирных кислот происходит из ацетил-КоА.
5.Для синтеза жирных кислот необходимы НАДФН, АТФ, Mn2+, биотин и СО2.
Синтез жирных кислот происходит в 3 этапа.
1) транспорт ацетил-КоА из митохондрий в цитозоль; 2) образование малонил-КоА; 3) удлинение жирной кислоты на 2 атома углерода за счет малонил-КоА до образования пальмитиновой кислоты.
1.Транспорт ацетил-КоАиз митохондрий в цитозоль осуществляется с помощью цитратного челночного механизма (рис.13.5 )

Рис. 10.5. Упрощенная схема цитратного челночного механизма и образования НАДФН
1.1. Цитратсинтаза катализирует реакцию взаимодействия ЩУК и ацетил-КоА с образованием цитрата
1.2. Цитрат траспортируется в цитозоль с помощью специфической транспортной системы.
1.3. В цитозоле цитрат взаимодействует с HS-KoA и под действием цитратлиазы и АТФ образуется ацетил-КоА и ЩУК.
1.4. ЩУК может вернуться в митохондрии с помощью транслоказы, но чаще восстанавливается до малата под действием НАД+-зависимой малатдегидрогеназы.
1.5. Малат декарбоксилируется НАДФ-зависимой малатдегидрогеназой (малик-фермент): Образующийся НАДФН+Н+ (50% потребности) используется для синтеза жирных кислот. Кроме этого генераторами НАДФН+Н+ (50%) являются пентозофосфатный путь и изоцитратдегидрогеназа.
1.6.Пируват транспортируетсяв митохондрии и под действием пируваткарбоксилазы образуется ЩУК.
2.Образование малонил-КоА.Ацетил-КоА карбоксилируется под действием ацетил-КоА-карбоксилазы.Это АТФ-зависимая реакция, для которой необходим витамин Н (биотин) и СО2.
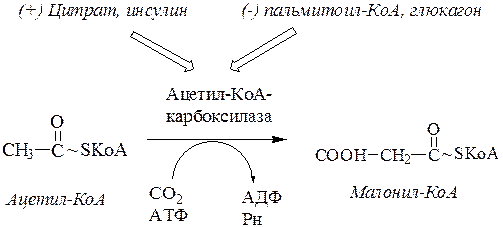
Эта реакция лимитирует скорость всего процесса синтеза жирных кислот: активаторы – цитрат и инсулин, ингибитор - синтезированная жирная кислота и глюкагон.
3.Удлинение жирной кислоты. Процесс протекает при участии мультиферментного синтазного комплекса. Он состоит из двух полипептидных цепей. Каждая полипептидная цепь содержит по 6 ферментов синтеза жирных кислот (трансацилаза, кетоацил-синтаза, кетоацил-редуктаза, гидратаза, еноил-редуктаза, тиоэстераза). Ферменты связаны между собой ковалентными связями. Ацилпереносящий белок (АПБ) является также частью полипептидной цепи, но это не фермент. Его функция связана только с переносом ацильных радикалов. В процессе синтеза важную роль играют SH-группы. Одна из них принадлежит 4-фосфопантетеину, входящему в состав АПБ и вторая - цистеину фермента кетоацил-синтазы. Первую называют центральной, а вторую периферической SH-группой.
Функциональная единица синтеза жирных кислот состоит из половины одного мономера, взаимодействующей с комплементарной половиной второго мономера. Следовательно, на синтазном комплексе одновременно синтезируются 2 жирные кислоты. Активной является только димерная форма комплекса. Перенос субстрата от фермента к ферменту происходит при участии АПБ.
Дата добавления: 2015-06-12; просмотров: 10514;