Физиологические свойства симпатической НС (СНС)
Благодаря мех-мам мультипликации в симпатических ганглиях распространяющиеся из них возбуждения широко охватывают сразу несколько органов. В результате разветвлённых влияний на иннервируемые органы СНС оказывает на них и на организм в целом генерализованные влияния (наиболее отчётливо прослеживается при эмоц. реакциях).
При возбуждении СНС усиливаются сердеч. сокращения, увелич. их ритм, повышается артер. давление, улучшается оксигенация тканей, усил. метаболизм, нарастает содержание глюкозы в крови, увелич. скорость проведения возбуждения в скелетных мышцах, повыш. их тонус, расширяются бронхи, увелич. объём лёгочной вентиляции, расшир. зрачки, увелич. секреция катехоламинов надпочечниками. Одновременно сниж. тонус пищеварит. тракта, ослаб. процессы всасывания и ферментативного расщепления в кишечнике.
СНС участвует в формир-нии агрессии, стресса, болевых реакций, мобилизует организм на борьбу, бегство, активное взаимодействие с окр. миром.
Хим. особенности. В снс передача с пре- на ганглионарные нейроны осуществ. с помощью ацетилхолина, а на эффекторы – посредством норадреналина. Исключение: постганглионарные сипатич. волокна к потовым железам и отдельным сосудам, в окончанииях которых выделяется ацетилхолин.
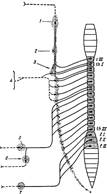 преганглионарные – сплошные линии, постганглионарные - прерывистые
преганглионарные – сплошные линии, постганглионарные - прерывистые
СНС представлена на периферии на значит. протяжении постганглионарными волокнами типа С, => возбуждение по СНС распростран. медленнее, чем по парасимпатической.
Эффекты снс более продолжительны, чем в парасимпат. нс.
Электрические потенциалы в симпатич. ганглиях характеризуются продолжительными следовыми явлениями.
При стимуляции преганглионарных волокон в симпатич. ганглиях сначала регистрируется быстрый возбуждающий постсинаптический потенциал (ВПСП), кот. сменяется пролонгированным медленным ТПСП. Вслед за этим снова регистрируется медленный ВПСП (длится минуты).
Начальная деполяризация обусловлена действием ацетилхолина на никотиновые рецепторы. Медленный ТПСП определяется действием дофамина, кот. секретируется малыми по размеру интернейронами ганглия. Интернейроны, в свою очередь, активизируются через мускариновые рецепторы.
Локализация преганглионарных и ганглионарных нейронов и иннервируемые ими органы. В отличие от парасимпат. нервов, которые выходят из различ. органов ЦНС, все симпат. нервы выходят из спинного мозга, иннервируют все органы и ткани организма. В последнем шейном и двух первых грудных сегментах спинного мозга расположен спиноцилиарный центр, иннервирующий гладкие мышцы глаза, в т.ч. мышцу, расширяющую зрачок. Эффекторные нейроны этого пути лежат в краниальном шейном симпат. ганглии. От 2-4 грудных сементов спин. мозга отходят симпат. волокна, иннервирующие слюнные железы. В первых трёх сегметах спин. мозга расположены эффектор. симпат. нейроны, иннервир. сердце. Ганглионарные клетки этого пути лежат в основном в звёздчатом ганглии или (реже) в узлах пограничного симпат. ствола. Сосуды и потовые железы иннервируются ото всех симпат. преганглионарных нейронов спин. мозга.
Больш-во симпат. симпат. ганглиев удалено от иннервируемых ими органов, поэтому от этих ганглиев идут длинные постганглионар. аксоны. Исключение – некоторые небольшие ганглии, располож. рядом с половыми органами.
Симпат. нервы регулируют функции всех органов и тканей организма, включая ЦНС и сенсорные рецепторы.
Медиаторы и рецепторы.
1. преганглионар. нейроны и рецепторы ганглионар. нейронов. Эфферентный вход в вегетативном ганглии (экстра- и интрогенный) представлен возбуждающим холинергическим преганглионаным волокном, образующим синапс с ганглионар. нейроном при помощи Н-холинорецепотра (медиатор – ацетилцелин). Рецептор получил своё название (Д. Ленги) из-за чувствительности к никотину: малые его дозы возуждают нейрон, малые – блокируют. В вегетативных ганглиях кроме него имеются различные нейропептиды (М-холинорецепторы, вставочные адренергические клетки, регулирующие возбудимость нейронов).
2. От ганглионарных нейронов отходят постганглионар. симпат. волокна, в окончаниях которых главным медиатором явл. норадреналин – около 90%. Он синтезируется из аминокислоты тирозина в аксоне ганглионар. нейрона, в основном в терминал. симпат. окончаниях, около 1% - в теле нейрона. В ответ на раздражение симпат. нейрона вместе с норадреналином выдел. также белок хромагрин дофамин-β-гидроксилаза, мет-энкефалин. Эту смесь дополняют медиаторы соседних нервных окончаний, гормоны, метаболиты и ионы эффекторных клеток.
3. Инактивация медиатора. 75-80% выделившегося в синаптическую щель норадреналина захватывается обратно пресинаптической мембраной и поступает в пузырьки. Не попавший туда норадреналин разрушается моноаминоксидазой (МАК), локализованной на пресинаптич. мембране. Часть выделившегося в синаптич. щель и не связавшегося с пост- и пресинаптич. рецепторами норадреналина разрушается в области рецепторов постсинаптич. Мембраны ферментом катехол-О-метилтран… феразой (КОМТ) клеточных эффекторов, другая – диффундирует в интерстиций… кровь. Вклад МАО и КОМТ в процессы инактивации выделившихся в синаптич. щель катехоламинов невелик, а доля участия этих ферментов в разных органах и тканях различна: в ЦНС действует в основном МАО, в периферич. симпат. НС – КОМТ.
4. Эффекторные рецепторы. Выделившийся норадреналин действует на α- и β-постсинаптич. адренорецепторы, являющиеся …гликопротедами. Это деление рецепторов основано на чувствительности их к различным фармакологическим препаратам: α-адренорецепторы блокируются фентоламином, а β- - пропранолом. Оба типа рецепторов дел. на 2 подтипа: α1 и α2, β1 и β2-адренорецепторы. В больш-ве органов, реагирующих на катехоламины, содержатся оба рецептора.
Связь постгаглионар. симпат. окончаний с адренорецепторами. Считается, что симпат. иннервацию имеют только α1 и β1-адренорецепторы, которые активизируются в основном медиатором норадреналином. α2- β2- преимущественно не имеют иннервации, расположены вне синапсов на клетках-эффекторах и активизируются циркулирующим в крови адреналином и диффундирующим норадреналином поле его выделения пресинаптич. окончаниями. α2- β2- обнаружены также на пресинаптич. рецепторах, где они выполняют регулирующую функцию. Прямая связь посредством синапса имеется в сердце, жировой ткани, сосудах и в ряде гладкомышечных органов (мышцы зрачка, матки, кишечник и т.д.) непрямой тип регуляции осуществляется в больш-ве органов: скеленая мускулатура, эндокрин. железы и т.д. В них медиатор из сосудистых симпат. сплетений диффундирует к окружающим сосуд клеткам либо разносится кроью. Другой вариант: симпат. нервные волокна образуют синаптич. контакты с нейронами инраорганной НС.
Эффекты активации α- β-адренорецепторов. В физиолог. условиях реакция какого-либо органа на адреналин и норадреналин, поступ. с кровью либо выделяющиеся при возбуждении симпат. нервов, зависит от преобладания α- или β-адрнорцптров в данном органе и различной их возбудимости.
Активация α1- и α2- мембран клеток-эффекторов ведёт к деполяризации их и ↑ активности органа, в т.ч. сфинктеров желудочно-кишечного тракта. Однако у миоцитов желудка и кишечника развив. гиперполяризация, при этом мускулатура органов расслабляется.
Если же симпат. ствол раздражается в грудной полости, то в больш-ве случаев регистрир-ся не тормозные, а стимуляторные реакции желудка и 12перстной кишки. Это осуществ. с помощью серотонинергических нервных волокон, имеющихся в составе симпат. стволов.
Активация β-адрнорцптров вызывает разнонаправленные эффекты в различных органах: в сердце возникает гиперполяризация, угнетение функции органа, расширение сосудов. При одновременной активации α- и β- сосуды сужаются следствие преобладания α-.
Механизм действия катехоламинов на эффектоные органы.
Во всех видах адрнпцпторов катехоламины взаимодействуют посредством G-белка. При этом возникают электрофизиоог. и биохим. процессы. При деполяризации наблюд. Усиление функции органа, при гиперполяризации – угнетение, например угнетение сокращений кишки. При активации обоих типов рецепторов возник. метаболические сдвиги с помощью вторых посредников (биохим. действия медиатора): при активации α- - инозитол-3-фосфата и Ca2+, β- - аденилатциклазы-цАМФ.
Чувствительность эффекторных клеток к медиатору может понижаться или повышаться.
Билет 42
Парасимпатическая часть автономной нервной системы имеет общую структуру, подобную симпатической части: здесь также выделяют центральные и периферические образования. Как и в симпатической части, передача возбуждения к исполнительному органу осуществляется по двухнейронному пути. Вместе с тем ряд признаков отличает парасимпатическую часть от симпатической.
Во-первых, центральные структуры парасимпатической части расположены в трех различных, далеко отстоящих друг от друга участках мозга; во-вторых, характерно наличие значительно более длинных преганглионарных и чрезвычайно коротких постганглионарных волокон; в-третьих, парасимпатические волокна иннервируют, как правило, только определенные зоны тела, которые также снабжаются симпатической, а в значительной части, кроме того, и метасимпатической иннервацией.
Центральные образования парасимпатической части автономной нервной системы включают ядра, лежащие в среднем, продолговатом и спинном мозге. В среднем мозге находится парасимпатическое добавочное ядро глазодвигательного нерва (ядро Якубовича, Вестфаля — Эдингера), расположенное вблизи передних бугров четверохолмия; в продолговатом мозге — три пары ядер, от которых начинаются преганглионарные волокна, выходящие из мозга в составе VII, IX, X пар черепных нервов (лицевого, языкоглоточного, блуждающего). Здесь проходят слюноотделительные, слезоотделительные, а также двигательный и секреторный пути для внутренних органов (блуждающий нерв). Парасимпатические ядра спинного мозга располагаются в области I—III или II—IV крестцовых сегментов в боковых рогах серого вещества.
Периферические структуры парасимпатической части автономной нервной системы включают нервные волокна и соответствующие ганглии. Преганглионарные волокна из среднего мозга выходят сбоку от ножек большого мозга в составе глазодвигательного нерва, проникают через глазную щель в глазницу и синаптически заканчиваются на эффекторных клетках расположенного в глубине глазницы ресничного узла. От него отходят два коротких ресничных нерва. Составляющие их постганглионарные волокна вступают в глазное яблоко, разветвляясь в аккомодационной мышце и сфинктере зрачка.
В продолговатом мозге нервные волокна из верхнего слюноотделительного ядра идут в составе лицевого нерва и, покидая его, образуют барабанную струну, которая позже присоединяется к язычному нерву. Последний достигает челюстного или подъязычного узла, постганглионарные волокна которого иннервируют подчелюстную слюнную железу. Преганглионарные волокна, выходящие из нижнего слюноотделительного ядра, вступают в языкоглоточный нерв и далее попадают в ушной узел. Его постганглионарные волокна являются секреторными для околоушной слюнной железы. Преганглионарные волокна из ядер слезоотделительного пути через лицевой нерв вступают в крылонебный узел, постганглионарные волокна которого достигают слюнной железы, желез слизистой оболочки носа и неба.
Блуждающий нерв является смешанным: он включает афферентные и эфферентные парасимпатические, чувствительные и двигательные соматические, а также эфферентные симпатические волокна. По выходе из черепа нерв образует два последовательно лежащих узла: верхний и нижний (яремный и узловой). Верхний узел содержит в основном чувствительные клетки, аналогичные клеткам спинномозговых узлов. От нижнего узла берут начало сердечный депрессорный нерв, возвратный гортанный нерв, пищеводные ветви. У корня легкого от блуждающего нерва отходят соответствующие веточки к легкому. В брюшной полости нерв переходит на желудок, формируя желудочное сплетение, от которого отходят стволики в чревное (солнечное) сплетение. Грудная и брюшная части блуждающего нерва могут рассматриваться лишь как проводники, связывающие центральные структуры с эффекторным аппаратом метасимпатической нервной системы.
Крестцовый отдел парасимпатической части нервной системы представлен тазовым нервом, который направляется к поверхности прямой кишки, где вместе с подчревным симпатическим нервом участвует в образовании тазового сплетения.
Дата добавления: 2015-06-05; просмотров: 1793;